Внутриклеточный механизм действия гормонов.
Гормоны, имеющие внутриклеточные рецепторы, свободно проникают через мембраны клеток-мишеней благодаря их высокой степени гидрофобности и липидорастворимости. Мембраны не являются барьерами для данного типа гормонов. Образующийся внутриклеточно ГР-комплекс диффузионным путем проникает через поры ядра и связывается с определенным участком ДНК, ответственным за присоединение того или иного гормона (HRE – hormone response element, см. табл. 3, рис. 5). В зависимости от того, какое влияние будет оказываться на транскрипцию генов ДНК (активация транскрипции или ее замедление), этот участок называется либо энхансером (от английского to enhance – усиливать), либо сайленсером (от английского silence – тишина). Каждый гормон-связывающий участок ДНК имеет консенсусную последовательность нуклеотидов (табл. 3), которая «узнается» ГР-комплексом, причем каждый консенсус состоит из двух шестинуклеотидных последовательностей, либо смежных, либо разделенных двумя или тремя нуклеотидами (N2 или N3).
Таблица 3
Примеры некоторых энхансерных участков ДНК, ответственных
За связывание ГР-комплексов гидрофобных гормонов
| ГР-комплекс | ДНК-последовательность |
| Андроген (тестостерон) | GG(A/T)ACAN2TGTTCT |
| Глюкокортикоиды (кортизол) | GGTACAN3TGTTCT |
| Йодтиронины (тироксин) | AGGTCAN3AGGTCA |
Конечным эффектом всех внутриклеточно действующих гормонов будет изменение количества мРНК, а, следовательно, и внутриклеточной концентрации какого-либо белка (или белков), что в результате приведет к определенному клеточному ответу (рост, дифференцировка, деление, выработка секретов, изменение метаболизма клетки и т. д.).
|
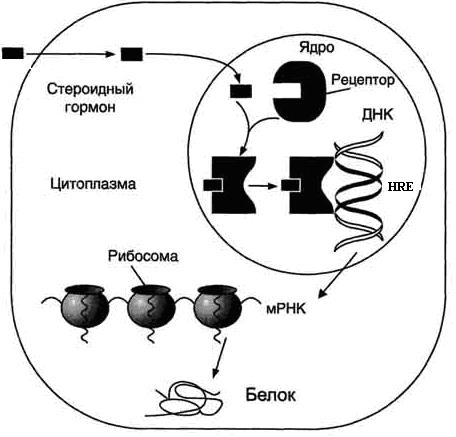


Рис. 5. Механизм регуляторного действия гормонов, имеющих внутриклеточные рецепторы (https://studfiles.net/preview/1219412/).
Например, мужские половые гормоны (андрогены), проникая в клетки мышечной ткани (миоциты), индуцируют транскрипцию генов ДНК, кодирующих аминокислотную последовательность основных сократительных белков (миозина, актина, тропонина, тропомиозина). Физиологическим ответом мышечной ткани на регуляторный сигнал гормонов данного типа будет увеличение массы, объема и сократительной способности мышечной ткани.
Женские половые гормоны (эстрогены) являются индукторами транскрипции генов белков клеток эндометрия матки, участвующих в его росте и пролиферации в определенную стадию овариально-менструального цикла.
Кортикостероидые гормоны класса глюкокортикоидов, действуя на гепатоциты, вызывают в них усиленную транскрипцию генов ферментов глюконеогенеза (пируваткарбоксилазы, фосфоенолпируваткарбоксикиназы, малатдегидрогеназы, фруктозо-1,6-дифосфатазы, глюкозо-6-фосфатазы). В результате физиологическим ответом гепатоцитов будет резкое увеличение поступления глюкозы из печени в кровь с развитием гиперглюкоземии.
При увеличении выработки минералокортикоидов (альдостерона) в клетках эпителия дистального отдела почечных канальцев наблюдается усиление транскрипции генов белков-ферментов, участвующих в реабсорбции ионов Na+, Cl- и HCO3-. Физиологический эффект такой регуляции представляет собой изменение ионного состава крови и мочи, а также изменение рН мочи и буферной емкости крови.
Регуляторное действие витамина D (1,25-диоксихолекальциферола) проявляется на клетках тонкого кишечника и канальцев почек как эффект усиления транскрипции генов ферментов, участвующих во всасывания кальция в кишечнике и реабсорбции ионов кальция из первичной мочи. Конечный результат суммации обоих этих эффектов – повышение уровня кальция в крови, что способствует последующей его утилизации в костной ткани.
Ретиноевая кислота, гормонально активное производное витамина А (ретинола), активирует транскрипцию генов ряда белков эпителиальных клеток (кожа, склера, роговица и др.), заставляя эти клетки дифференцироваться в более зрелые формы. Дефицит витамина А приводит не только к нарушениям зрения, но и к развитию поражений эпителиальных покровов кожи и глаз.
Разумеется, перечисленные примеры не исчерпывают всего спектра регуляторного действия указанных гормонов, а лишь иллюстрируют принципиально одинаковый механизм развития конечных физиологических ответов клеток-мишеней на действие гормонов с внутриклеточным типом рецепции.
В целом, регуляторный эффект гормонов, действующих через внутриклеточные рецепторы, может быть охарактеризован как экстенсивная регуляция, т. е. регуляция через изменение количества (концентрации) белков (в частности белков-ферментов) в клетках-мишенях (рис. 5). Гормоны, действующие через мембранные рецепторы, характеризуются интенсивным характером регуляторного действия. Их эффект не вызывает изменения количества белков и ферментов в клетках-мишенях, а способствует изменению активности ферментов, имеющихся в этих клетках (см. следующий раздел).