Клетка
Клетка, элементарная единица живого. Клетка отграничена от других клеток или от внешней среды специальной мембраной и имеет ядро или его эквивалент, в котором сосредоточена основная часть химической информации, контролирующей наследственность. Изучением строения клетки занимается цитология, функционированием – физиология. Наука, изучающая состоящие из клеток ткани, называется гистологией.
Существуют одноклеточные организмы, тело которых целиком состоит из одной клетки. К этой группе относятся бактерии и протисты (простейшие животные и одноклеточные водоросли). Иногда их также называют бесклеточными, но термин одноклеточные употребляется чаще. Настоящие многоклеточные животные (Metazoa) и растения (Metaphyta) содержат множество клеток.
Абсолютное большинство тканей состоит из клеток, однако имеются и некоторые исключения. Тело слизевиков (миксомицетов), например, состоит из однородной, не разделенной на клетки субстанции с многочисленными ядрами. Сходным образом организованы и некоторые животные ткани, в частности сердечная мышца. Вегетативное тело (таллом) грибов образовано микроскопическими нитями – гифами, нередко сегментированными; каждая такая нить может считаться эквивалентом клетки, хотя и нетипичной формы.
Некоторые не участвующие в метаболизме структуры тела, в частности раковины, жемчужины или минеральная основа костей, образованы не клетками, а продуктами их секреции. Другие, например древесина, кора, рога, волосы и наружный слой кожи, – не секреторного происхождения, а образованы из мертвых клеток.
Мелкие организмы, такие, как коловратки, состоят всего из нескольких сотен клеток. Для сравнения: в человеческом организме насчитывается ок. 1014 клеток, в нем каждую секунду погибают и замещаются новыми 3 млн. эритроцитов, и это всего одна десятимиллионная часть от общего количества клеток тела.
Обычно размеры растительных и животных клеток колеблются в пределах от 5 до 20 мкм в поперечнике. Типичная бактериальная клетка значительно меньше – ок. 2 мкм, а наименьшая из известных – 0,2 мкм.
Некоторые свободноживущие клетки, например такие простейшие, как фораминиферы, могут достигать нескольких сантиметров; они всегда имеют много ядер. Клетки тонких растительных волокон достигают в длину одного метра, а отростки нервных клеток достигают у крупных животных нескольких метров. При такой длине объем этих клеток небольшой, а поверхность очень велика.
Самые крупные клетки – это неоплодотворенные яйца птиц, заполненные желтком. Наибольшее яйцо (и, следовательно, наибольшая клетка) принадлежало вымершей громадной птице – эпиорнису (Aepyornis). Предположительно его желток весил ок. 3,5 кг. Самое крупное яйцо у ныне живущих видов принадлежит страусу, его желток весит ок. 0,5 кг.
Как правило, клетки крупных животных и растений лишь немногим больше клеток мелких организмов. Слон больше мыши не потому, что его клетки крупнее, а в основном потому, что самих клеток значительно больше. Существуют группы животных, например коловратки и нематоды, у которых количество клеток в организме остается постоянным. Таким образом, хотя крупные виды нематод имеют большее количество клеток, чем мелкие, основное различие в размерах обусловлено в этом случае все же большими размерами клеток.
В пределах данного типа клеток их размеры обычно зависят от плоидности, т.е. от числа наборов хромосом, присутствующих в ядре. Тетраплоидные клетки (с четырьмя наборами хромосом) в 2 раза больше по объему, чем диплоидные клетки (с двойным набором хромосом). Плоидность растения можно увеличить путем введения в него растительного препарата колхицина. Поскольку подвергнутые такому воздействию растения имеют более крупные клетки, они и сами крупнее. Однако это явление можно наблюдать только на полиплоидах недавнего происхождения. У эволюционно древних полиплоидных растений размеры клеток подвержены «обратной регуляции» в сторону нормальных величин несмотря на увеличение числа хромосом.
Структура клетки
Одно время клетка рассматривалась как более или менее гомогенная капелька органического вещества, которую называли протоплазмой или живой субстанцией. Этот термин устарел после того, как выяснилось, что клетка состоит из множества четко обособленных структур, получивших название клеточных органелл («маленьких органов»).
Химический состав. Обычно 70–80 % массы клетки составляет вода, в которой растворены разнообразные соли и низкомолекулярные органические соединения. Наиболее характерные компоненты клетки – белки и нуклеиновые кислоты. Некоторые белки являются структурными компонентами клетки, другие – ферментами, т.е. катализаторами, определяющими скорость и направление протекающих в клетках химических реакций. Нуклеиновые кислоты служат носителями наследственной информации, которая реализуется в процессе внутриклеточного синтеза белков.
Часто клетки содержат некоторое количество запасных веществ, служащих пищевым резервом. Растительные клетки в основном запасают крахмал – полимерную форму углеводов. В клетках печени и мышц запасается другой углеводный полимер – гликоген. К часто запасаемым продуктам относится также жир, хотя некоторые жиры выполняют иную функцию, а именно служат важнейшими структурными компонентами. Белки в клетках (за исключением клеток семян) обычно не запасаются.
Описать типичный состав клетки не представляется возможным прежде всего потому, что существуют большие различия в количестве запасаемых продуктов и воды. В клетках печени содержится, например, 70% воды, 17% белков, 5% жиров, 2% углеводов и 0,1% нуклеиновых кислот; оставшиеся 6% приходятся на соли и низкомолекулярные органические соединения, в частности аминокислоты. Растительные клетки обычно содержат меньше белков, значительно больше углеводов и несколько больше воды; исключение составляют клетки, находящиеся в состоянии покоя. Покоящаяся клетка пшеничного зерна, являющегося источником питательных веществ для зародыша, содержит ок. 12% белков (в основном это запасаемый белок), 2% жиров и 72% углеводов. Количество воды достигает нормального уровня (70–80%) только в начале прорастания зерна.
Главные части клетки. Некоторые клетки, в основном растительные и бактериальные, имеют наружную клеточную стенку. У высших растений она состоит из целлюлозы. Стенка окружает собственно клетку, защищая ее от механических воздействий. Клетки, в особенности бактериальные, могут также секретировать слизистые вещества, образуя тем самым вокруг себя капсулу, которая, как и клеточная стенка, выполняет защитную функцию.
Именно с разрушением клеточных стенок связана гибель многих бактерий под действием пенициллина. Дело в том, что внутри бактериальной клетки концентрация солей и низкомолекулярных соединений очень высока, а потому в отсутствие укрепляющей стенки вызванный осмотическим давлением приток воды в клетку может привести к ее разрыву. Пенициллин, препятствующий во время роста клетки формированию ее стенки, как раз и приводит к разрыву (лизису) клетки.
Клеточные стенки и капсулы не участвуют в метаболизме, и часто их удается отделить, не убивая клетку. Таким образом, их можно считать наружными вспомогательными частями клетки. У клеток животных клеточные стенки и капсулы, как правило, отсутствуют.
Собственно клетка состоит из трех основных частей. Под клеточной стенкой, если она имеется, находится клеточная мембрана. Мембрана окружает гетерогенный материал, называемый цитоплазмой. В цитоплазму погружено круглое или овальное ядро. Ниже мы рассмотрим более подробно структуру и функции этих частей клетки.
Клеточная мембрана
Клеточная мембрана – очень важная часть клетки. Она удерживает вместе все клеточные компоненты и разграничивает внутреннюю и наружную среду. Кроме того, модифицированные складки клеточной мембраны образуют многие органеллы клетки.
Клеточная мембрана представляет собой двойной слой молекул (бимолекулярный слой, или бислой). В основном это молекулы фосфолипидов и других близких к ним веществ. Липидные молекулы имеют двойственную природу, проявляющуюся в том, как они ведут себя по отношению к воде. Головы молекул гидрофильные, т.е. обладают сродством к воде, а их углеводородные хвосты гидрофобны. Поэтому при смешивании с водой липиды образуют на ее поверхности пленку, аналогичную пленке масла; при этом все их молекулы ориентированы одинаково: головы молекул – в воде, а углеводородные хвосты – над ее поверхностью.
В клеточной мембране два таких слоя, и в каждом из них головы молекул обращены наружу, а хвосты – внутрь мембраны, один к другому, не соприкасаясь таким образом с водой. Толщина такой мембраны ок. 7 нм. Кроме основных липидных компонентов, она содержит крупные белковые молекулы, которые способны «плавать» в липидном бислое и расположены так, что одна их сторона обращена внутрь клетки, а другая соприкасается с внешней средой. Некоторые белки находятся только на наружной или только на внутренней поверхности мембраны или лишь частично погружены в липидный бислой.
Основная функция клеточной мембраны заключается в регуляции переноса веществ в клетку и из клетки. Поскольку мембрана физически в какой-то мере похожа на масло, вещества, растворимые в масле или в органических растворителях, например эфир, легко проходят сквозь нее. То же относится и к таким газам, как кислород и диоксид углерода. В то же время мембрана практически непроницаема для большинства водорастворимых веществ, в частности для сахаров и солей. Благодаря этим свойствам она способна поддерживать внутри клетки химическую среду, отличающуюся от наружной. Например, в крови концентрация ионов натрия высокая, а ионов калия – низкая, тогда как во внутриклеточной жидкости эти ионы присутствуют в обратном соотношении. Аналогичная ситуация характерна и для многих других химических соединений.
Очевидно, что клетка тем не менее не может быть полностью изолирована от окружающей среды, так как должна получать вещества, необходимые для метаболизма, и избавляться от его конечных продуктов. К тому же липидный бислой не является полностью непроницаемым даже для водорастворимых веществ, а пронизывающие его т.н. «каналообразующие» белки создают поры, или каналы, которые могут открываться и закрываться (в зависимости от изменения конформации белка) и в открытом состоянии проводят определенные иона (Na+, K+, Ca2+) по градиенту концентрации. Следовательно, разница концентраций внутри клетки и снаружи не может поддерживаться исключительно за счет малой проницаемости мембраны. На самом деле в ней имеются белки, выполняющие функцию молекулярного «насоса»: они транспортируют некоторые вещества как внутрь клетки, так и из нее, работая против градиента концентрации. В результате, когда концентрация, например, аминокислот внутри клетки высокая, а снаружи низкая, аминокислоты могут тем не менее поступать из внешней среды во внутреннюю. Такой перенос называется активным транспортом, и на него затрачивается энергия, поставляемая метаболизмом. Мембранные насосы высокоспецифичны: каждый из них способен транспортировать либо только ионы определенного металла, либо аминокислоту, либо сахар. Специфичны также и мембранные ионные каналы.
Такая избирательная проницаемость физиологически очень важна, и ее отсутствие – первое свидетельство гибели клетки. Это легко проиллюстрировать на примере свеклы. Если живой корень свеклы погрузить в холодную воду, то он сохраняет свой пигмент; если же свеклу кипятить, то клетки погибают, становятся легко проницаемыми и теряют пигмент, который и окрашивает воду в красный цвет.
Крупные молекулы типа белковых клетка может «заглатывать». Под влиянием некоторых белков, если они присутствуют в жидкости, окружающей клетку, в клеточной мембране возникает впячивание, которое затем смыкается, образуя пузырек – небольшую вакуоль, содержащую воду и белковые молекулы; после этого мембрана вокруг вакуоли разрывается, и содержимое попадает внутрь клетки. Такой процесс называется пиноцитозом (буквально «питье клетки»), или эндоцитозом.
Более крупные частички, например частички пищи, могут поглощаться аналогичным образом в ходе т.н. фагоцитоза. Как правило, вакуоль, образующаяся при фагоцитозе, крупнее, и пища переваривается ферментами лизосом внутри вакуоли до разрыва окружающей ее мембраны. Такой тип питания характерен для простейших, например для амеб, поедающих бактерий. Однако способность к фагоцитозу свойственна и клеткам кишечника низших животных, и фагоцитам – одному из видов белых кровяных клеток (лейкоцитов) позвоночных. В последнем случае смысл этого процесса заключается не в питании самих фагоцитов, а в разрушении ими бактерий, вирусов и другого инородного материала, вредного для организма.
Функции вакуолей могут быть и другими. Например, простейшие, живущие в пресной воде, испытывают постоянный осмотический приток воды, так как концентрация солей внутри клетки гораздо выше, чем снаружи. Они способны выделять воду в специальную экскретирующую (сократительную) вакуоль, которая периодически выталкивает свое содержимое наружу.
В растительных клетках часто имеется одна большая центральная вакуоль, занимающая почти всю клетку; цитоплазма при этом образует лишь очень тонкий слой между клеточной стенкой и вакуолью. Одна из функций такой вакуоли – накопление воды, позволяющее клетке быстро увеличиваться в размерах. Эта способность особенно необходима в период, когда растительные ткани растут и образуют волокнистые структуры.
В тканях в местах плотного соединения клеток их мембраны содержат многочисленные поры, образованные пронизывающими мембрану белками – т.н. коннексонами. Поры прилежащих клеток располагаются друг против друга, так что низкомолекулярные вещества могут перегодить из клетки в клетку – эта химическая система коммуникации координирует их жизнедеятельность. Один из примеров такой координации – наблюдаемое во многих тканях более или менее синхронное деление соседних клеток.
Цитоплазма
В цитоплазме имеются внутренние мембраны, сходные с наружной и образующие органеллы различного типа. Эти мембраны можно рассматривать как складки наружной мембраны; иногда внутренние мембраны составляют единое целое с наружной, но часто внутренняя складка отшнуровывается, и контакт с наружной мембраной прерывается. Однако даже в случае сохранения контакта внутренняя и наружная мембраны не всегда химически идентичны. В особенности различается состав мембранных белков в разных клеточных органеллах.
Эндоплазматический ретикулум. Состоящая из канальцев и пузырьков сеть внутренних мембран тянется от поверхности клетки до ядра. Эта сеть называется эндоплазматическим ретикулумом. Часто отмечалось, что канальцы открываются на поверхности клетки, и эндоплазматический ретикулум, таким образом, играет роль микроциркуляторного аппарата, через который внешняя среда может непосредственно взаимодействовать со всем содержимым клетки. Такое взаимодействие было обнаружено в некоторых клетках, в частности в мышечных, но пока не ясно, является ли оно универсальным. Во всяком случае транспорт ряда веществ по этим канальцам из одной части клетки в другую действительно происходит.
Крошечные тельца, называемые рибосомами, покрывают поверхность эндоплазматического ретикулума, особенно вблизи ядра. Диаметр рибосом ок. 15 нм, они состоят наполовину из белков, наполовину из рибонуклеиновых кислот. Их основная функция – синтез белков; к их поверхности прикрепляются матричная (информационная) РНК и аминокислоты, связанные с транспортными РНК. Участки ретикулума, покрытые рибосомами, называют шероховатым эндоплазматическим ретикулумом, а лишенные их – гладким. Кроме рибосом, на эндоплазматическом ретикулуме адсорбированы или иным образом к нему присоединены различные ферменты, в том числе системы ферментов, обеспечивающих использование кислорода для образования стеролов и для обезвреживания некоторых ядов. В неблагоприятных условиях эндоплазматический ретикулум быстро дегенерирует, и поэтому его состояние служит чувствительным индикатором здоровья клетки.
Аппарат Гольджи. Аппарат Гольджи (комплекс Гольджи) – это специализированная часть эндоплазматического ретикулума, состоящая из собранных в стопки плоских мембранных мешочков. Он участвует в секреции клеткой белков (в нем происходит упаковка секретируемых белков в гранулы) и поэтому особенно развит в клетках, выполняющих секреторную функцию. К важным функциям аппарата Гольджи относится также присоединение углеводных групп к белкам и использование этих белков для построения клеточной мембраны и мембраны лизосом. У некоторых водорослей в аппарате Гольджи осуществляется синтез волокон целлюлозы.
Лизосомы – это маленькие, окруженные одинарной мембраной пузырьки. Они отпочковываются от аппарата Гольджи и, возможно, от эндоплазматического ретикулума. Лизосомы содержат разнообразные ферменты, которые расщепляют крупные молекулы, в частности белковые. Из-за своего разрушительного действия эти ферменты как бы «заперты» в лизосомах и высвобождаются только по мере надобности. Так, при внутриклеточном пищеварении ферменты выделяются из лизосом в пищеварительные вакуоли. Лизосомы бывают необходимы и для разрушения клеток; например, во время превращения головастика во взрослую лягушку высвобождение лизосомных ферментов обеспечивает разрушение клеток хвоста. В данном случае это нормально и полезно для организма, но иногда такое разрушение клеток носит патологический характер. Например, при вдыхании асбестовой пыли она может проникнуть в клетки легких, и тогда происходит разрыв лизосом, разрушение клеток и развивается легочное заболевание.
Митохондрии и хлоропласты. Митохондрии – относительно крупные мешковидные образования с довольно сложной структурой. Они состоят из матрикса, окруженного внутренней мембраной, межмембранного пространства и наружной мембраны. Внутренняя мембрана сложена в складки, называемые кристами. На кристах размещаются скопления белков. Многие из них – ферменты, катализирующие окисление продуктов распада углеводов; другие катализируют реакции синтеза и окисления жиров. Вспомогательные ферменты, участвующие в этих процессах, растворены в матриксе митохондрий.
В митохондриях протекает окисление органических веществ, сопряженное с синтезом аденозинтрифосфата (АТФ). Распад АТФ с образованием аденозиндифосфата (АДФ) сопровождается выделением энергии, которая расходуется на различные процессы жизнедеятельности, например на синтез белков и нуклеиновых кислот, транспорт веществ внутрь клетки и из нее, передачу нервных импульсов или мышечное сокращение. Митохондрии, таким образом, являются энергетическими станциями, перерабатывающими «топливо» – жиры и углеводы – в такую форму энергии, которая может быть использована клеткой, а следовательно, и организмом в целом.
Растительные клетки тоже содержат митохондрии, но основной источник энергии для yих клеток – свет. Световая энергия используется этими клетками для образования АТФ и синтеза углеводов из диоксида углерода и воды. Хлорофилл – пигмент, аккумулирующий световую энергию, – находится в хлоропластах. Хлоропласты, подобно митохондриям, имеют внутреннюю и наружную мембраны. Из выростов внутренней мембраны в процессе развития хлоропластов возникают т.н. тилакоидные мембраны; последние образуют уплощенные мешочки, собранные в стопки наподобие столбика монет; эти стопки, называемые гранами, содержат хлорофилл. Кроме хлорофилла, в хлоропластах имеются и все другие компоненты, необходимые для фотосинтеза.
Некоторые специализированные хлоропласты не осуществляют фотосинтез, а несут другие функции, например обеспечивают запасание крахмала или пигментов.
Относительная автономия. В некоторых отношениях митохондрии и хлоропласты ведут себя как автономные организмы. Например, так же, как и сами клетки, которые возникают только из клеток, митохондрии и хлоропласты образуются только из предсуществующих митохондрий и хлоропластов. Это было продемонстрировано в опытах на растительных клетках, у которых образование хлоропластов подавляли антибиотиком стрептомицином, и на клетках дрожжей, где образование митохондрий подавляли другими препаратами. После таких воздействий клетки уже никогда не восстанавливали отсутствующие органеллы. Причина в том, что митохондрии и хлоропласты содержат определенное количество собственного генетического материала (ДНК), который кодирует часть их структуры. Если эта ДНК утрачивается, что и происходит при подавлении образования органелл, то структура не может быть воссоздана. Оба типа органелл имеют свою собственную белок-синтезирующую систему (рибосомы и транспортные РНК), которая несколько отличается от основной белок-синтезирующей системы клетки; известно, например, что белок-синтезирующая система органелл может быть подавлена с помощью антибиотиков, тогда как на основную систему они не действуют.
ДНК органелл ответственна за основную часть внехромосомной, или цитоплазматической, наследственности. Внехромосомная наследственность не подчиняется менделевским законам, так как при делении клетки ДНК органелл передается дочерним клеткам иным путем, нежели хромосомы. Изучение мутаций, которые происходят в ДНК органелл и ДНК хромосом, показало, что ДНК органелл отвечает лишь за малую часть структуры органелл; большинство их белков закодированы в генах, расположенных в хромосомах.
Частичная генетическая автономия рассматриваемых органелл и особенности их белок-синтезирующих систем послужили основой для предположения, что митохондрии и хлоропласты произошли от симбиотических бактерий, которые поселились в клетках 1–2 млрд. лет назад. Современным примером такого симбиоза могут служить мелкие фотосинтезирующие водjросли, которые живут внутри клеток некоторых кораллов и моллюсков. Водоросли обеспечивают своих хозяев кислородом, а от них получают питательные вещества.
Фибриллярные структуры. Цитоплазма клетки представляет собой вязкую жидкость, поэтому можно ожидать, что из-за поверхностного натяжения клетка должна иметь сферическую форму, за исключением тех случаев, когда клетки плотно упакованы. Однако обычно этого не наблюдается. Многие простейшие имеют плотные покровы или оболочки, которые придают клетке определенную, несферическую форму. Тем не менее даже без оболочки клетки могут поддерживать несферическую форму из-за того, что цитоплазма структурируется с помощью многочисленных, довольно жестких, параллельно расположенных волокон. Последние образованы полыми микротрубочками, которые состоят из белковых единиц, организованных в виде спирали.
Некоторые простейшие образуют псевдоподии – длинные тонкие цитоплазматические выросты, которыми они захватывают пищу. Псевдоподии сохраняют свою форму благодаря жесткости микротрубочек. Если гидростатическое давление возрастает примерно до 100 атмосфер, микротрубочки распадаются и клетка приобретает форму капли. Когда же давление возвращается к норме, вновь идет сборка микротрубочек и клетка образует псевдоподии. Сходным образом на изменение давления реагируют и многие другие клетки, что подверждает участие микротрубочек в сохранении формы клетки. Сборка и распад микротрубочек, необходимые для того, чтобы клетка могла быстро менять форму, происходят и в отсутствие изменений давления.
Из микротрубочек формируются также фибриллярные структуры, служащие органами движения клетки. У некоторых клеток имеются бичевидные выросты, называемые жгутиками, или же реснички – их биение обеспечивает движение клетки в воде. Если клетка неподвижна, эти структуры гонят воду, частицы пищи и другие частицы к клетке или от клетки. Жгутики относительно крупные, и обычно клетка имеет только один, изредка несколько жгутиков. Реснички гораздо мельче и покрывают всю поверхность клетки. Хотя эти структуры свойственны главным образом простейшим, они могут присутствовать и у высокоорганизованных форм. В человеческом организме ресничками выстланы все дыхательные пути. Попадающие в них небольшие частички обычно улавливаются слизью на клеточной поверхности, и реснички продвигают их вместе со слизью наружу, защищая таким образом легкие. Мужские половые клетки большинства животных и некоторых низших растений движутся с помощью жгутика.
Существуют и другие типы клеточного движения. Один из них – амебоидное движение. Амеба, а также некоторые клетки многоклеточных организмов «перетекают» с места на место, т.е. движутся за счет тока содержимого клетки. Постоянный ток вещества существует и внутри растительных клеток, однако он не влечет за собой передвижения клетки в целом. Наиболее изученный тип клеточного движения – сокращение мышечных клеток; оно осуществляется путем скольжения фибрилл (белковых нитей) относительно друг друга, что приводит к укорочению клетки.
Ядро
Ядро окружено двойной мембраной. Очень узкое (порядка 40 нм) пространство между двумя мембранами называется перинуклеарным. Мембраны ядра переходят в мембраны эндоплазматического ретикулума, а перинуклеарное пространство открывается в ретикулярное. Обычно ядерная мембрана имеет очень узкие поры. По-видимому, через них осуществляется перенос крупных молекул, таких, как информационная РНК, которая синтезируется на ДНК, а затем поступает в цитоплазму.
Основная часть генетического материала находится в хромосомах клеточного ядра. Хромосомы состоят из длинных цепей двуспиральной ДНК, к которой прикрепляются основные (т.е. обладающие щелочными свойствами) белки. Иногда в хромосомах имеется несколько идентичных цепей ДНК, лежащих рядом друг с другом, – такие хромосомы называются политенными (многонитчатыми). Число хромосом у разных видов неодинаково. Диплоидные клетки тела человека содержат 46 хромосом, или 23 пары.
В неделящейся клетке хромосомы прикреплены в одной или нескольких точках к ядерной мембране. В обычном неспирализованном состоянии хромосомы настолько тонки, что не видны в световой микроскоп. На определенных локусах (участках) одной или нескольких хромосом формируется присутствующее в ядрах большинства клеток плотное тельце – т.н. ядрышко. В ядрышках происходит синтез и накопление РНК, используемой для построения рибосом, а также некоторых других типов РНК.
Деление клетки
Хотя все клетки появляются путем деления предшествующей клетки, не все они продолжают делиться. Например, нервные клетки мозга, однажды возникнув, уже не делятся. Их количество постепенно уменьшается; поврежденные ткани мозга не способны восстанавливаться путем регенерации. Если же клетки продолжают делиться, то им свойствен клеточный цикл, состоящий из двух основных стадий: интерфазы и митоза.
Сама интерфаза состоит из трех фаз: G1, S и G2. Ниже указана их продолжительность, типичная для растительных и животных клеток.
G1 (4–8 ч). Это фаза начинается сразу после рождения клетки. На протяжении фазы G1 клетка, за исключением хромосом (которые не изменяются), увеличивает свою массу. Если клетка в дальнейшем не делится, то остается в этой фазе.
S (6–9 ч). Масса клетки продолжает увеличиваться, и происходит удвоение (дупликация) хромосомной ДНК. Тем не менее хромосомы остаются одинарными по структуре, хотя и удвоенными по массе, так как две копии каждой хромосомы (хроматиды) все еще соединены друг с другом по всей длине.
G2. Масса клетки продолжает увеличиваться до тех пор, пока она приблизительно вдвое не превысит начальную, а затем наступает митоз.
МИТОЗ
После того как хромосомы удвоились, каждая из дочерних клеток должна получить полный набор хромосом. Простое деление клетки не может этого обеспечить – такой результат достигается посредством процесса, называемого митозом. Если не вдаваться в детали, то началом этого процесса следует считать выстраивание хромосом в экваториальной плоскости клетки. Затем каждая хромосома продольно расщепляется на две хроматиды, которые начинают расходиться в противоположных направлениях, становясь самостоятельными хромосомами. В итоге на двух концах клетки располагается по полному набору хромосом. Далее клетка делится на две, и каждая дочерняя клетка получает полный набор хромосом.
Ниже приводится описание митоза в типичной животной клетке. Его принято разделять на четыре стадии.
I. Профаза. Особая клеточная структура – центриоль – удваивается (иногда это удвоение происходит в S-периоде интерфазы), и две центриоли начинают расходиться к противоположным полюсам ядра. Ядерная мембрана разрушается; одновременно специальные белки объединяются (агрегируют), формируя микротрубочки в виде нитей. Центриоли, расположенные теперь на противоположных полюсах клетки, оказывают организующее воздействие на микротрубочки, которые в результате выстраиваются радиально, образуя структуру, напоминающую по внешнему виду цветок астры («звезда»). Другие нити из микротрубочек протягиваются от одной центриоли к другой, образуя т.н. веретено деления. В это время хромосомы находятся в спирализованном состоянии, напоминая пружину. Они хорошо видны в световом микроскопе, особенно после окрашивания. В профазе хромосомы расщепляются, но хроматиды все еще остаются скрепленными попарно в зоне центромеры – хромосомной органеллы, сходной по функциям с центриолью. Центромеры тоже оказывают организующее воздействие на нити веретена, которые теперь тянутся от центриоли к центромере и от нее к другой центриоли.
II. Метафаза. Хромосомы, до этого момента расположенные беспорядочно, начинают двигаться, как бы влекомые нитями веретена, прикрепленными к их центромерам, и постепенно выстраиваются в одной плоскости в определенном положении и на равном расстоянии от обоих полюсов. Лежащие в одной плоскости центромеры вместе с хромосомами образуют т.н. экваториальную пластинку. Центромеры, соединяющие пары хроматид, делятся, после чего сестринские хромосомы полностью разъединяются.
III. Анафаза. Хромосомы каждой пары движутся в противоположных направлениях к полюсам, их как бы тащат нити веретена. При этом образуются нити и между центромерами парных хромосом.
IV. Телофаза. Как только хромосомы приближаются к противоположным полюсам, сама клетка начинает делиться вдоль плоскости, в которой находилась экваториальная пластинка. В итоге образуются две клетки. Нити веретена разрушаются, хромосомы раскручиваются и становятся невидимыми, вокруг них формируется ядерная мембрана. Клетки возвращаются в фазу G1 интерфазы. Весь процесс митоза занимает около часа.
Детали митоза несколько варьируют в разных типах клеток. В типичной растительной клетке образуется веретено, но отсутствуют центриоли. У грибов митоз происходит внутри ядра, без предшествующего распада ядерной мембраны.
Деление самой клетки, называемое цитокинезом, не имеет жесткой связи с митозом. Иногда один или несколько митозов проходят без клеточного деления; в результате образуются многоядерные клетки, часто встречающиеся у водорослей. Если из яйцеклетки морского ежа удалить путем микроманипуляций ядро, то веретено после этого продолжает формироваться и яйцеклетка продолжает делиться. Это показывает, что наличие хромосом не является необходимым условием для деления клетки.
Размножение с помощью митоза называют бесполым размножением, вегетативным размножением или клонированием. Его наиболее важный аспект – генетический: при таком размножении не происходит расхождения наследственных факторов у потомства. Образующиеся дочерние клетки генетически в точности такие же, как и материнская. Митоз – это единственный способ самовоспроизведения у видов, не имеющих полового размножения, например у многих одноклеточных. Тем не менее даже у видов с половым размножением клетки тела делятся посредством митоза и происходят от одной клетки – оплодотворенного яйца, а потому все они генетически идентичны. Высшие растения могут размножаться бесполым путем (с помощью митоза) саженцами и усами (известный пример – клубника).
Мейоз
Половое размножение организмов осуществляется с помощью специализированных клеток, т.н. гамет, – яйцеклетки (яйца) и спермия (сперматозоида). Гаметы, сливаясь, образуют одну клетку – зиготу. Каждая гамета гаплоидна, т.е. имеет по одному набору хромосом. Внутри набора все хромосомы разные, однако каждой хромосоме яйцеклетки соответствует одна из хромосом спермия. Зигота, таким образом, содержит уже пару таких соответствующих друг другу хромосом, которые называют гомологичными. Гомологичные хромосомы сходны, поскольку имеют одни и те же гены или их варианты (аллели), определяющие специфические признаки. Например, одна из парных хромосом может иметь ген, кодирующий группу крови А, а другая – его вариант, кодирующий группу крови В. Хромосомы зиготы, происходящие из яйцеклетки, являются материнскими, а происходящие из спермия – отцовскими.
В результате многократных митотических делений из образовавшейся зиготы возникает либо многоклеточный организм, либо многочисленные свободноживущие клетки, как это происходит у обладающих половым размножением простейших и у одноклеточных водорослей.
При образовании гамет диплоидный набор хромосом, имевшийся у зиготы, должен наполовину уменьшиться (редуцироваться). Если бы этого не происходило, то в каждом поколении слияние гамет приводило бы к удвоению набора хромосом. Редукция до гаплоидного числа хромосом происходит в результате редукционного деления – т.н. мейоза, который представляет собой вариант митоза.
Расщепление и рекомбинация. Особенность мейоза состоит в том, что при клеточном делении экваториальную пластинку образуют пары гомологичных хромосом, а не удвоенные индивидуальные хромосомы, как при митозе. Парные хромосомы, каждая из которых осталась одинарной, расходятся к противоположным полюсам клетки, клетка делится, и в результате дочерние клетки получают половинный, по сравнению с зиготой, набор хромосом.
Для примера предположим, что гаплоидный набор состоит из двух хромосом. В зиготе (и соответственно во всех клетках организма, продуцирующего гаметы) присутствуют материнские хромосомы А и В и отцовские А' и В'. Во время мейоза они могут разделиться следующим образом:
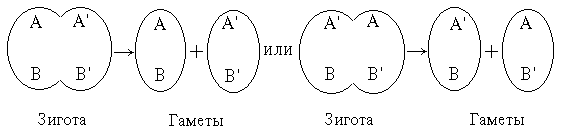
Наиболее важен в этом примере тот факт, что при расхождении хромосом вовсе не обязательно образуется исходный материнский и отцовский набор, а возможна рекомбинация генов, как в гаметах АВ' и А'В в приведенной схеме.
Теперь предположим, что пара хромосом АА' содержит два аллеля – a и b – гена, определяющего группы крови А и В. Сходным образом пара хромосом ВВ' содержит аллели m и n другого гена, определяющего группы крови M и N. Разделение этих аллелей может идти следующим образом:

Очевидно, что получившиеся гаметы могут содержать любую из следующих комбинаций аллелей двух генов: am, bn, bm или an.
Если имеется большее число хромосом, то пары аллелей будут расщепляться независимо по тому же принципу. Это означает, что одни и те же зиготы могут продуцировать гаметы с различными комбинациями аллелей генов и давать начало разным генотипам в потомстве.
Мейотическое деление. Оба приведенных примера иллюстрируют принцип мейоза. На самом деле мейоз – значительно более сложный процесс, так как включает два последовательных деления. Главное в мейозе то, что хромосомы удваиваются только один раз, тогда как клетка делится дважды, в результате чего происходит редукция числа хромосом и диплоидный набор превращается в гаплоидный.
Во время профазы первого деления гомологичные хромосомы конъюгируют, т. е. сближаются попарно. В результате этого очень точного процесса каждый ген оказывается напротив своего гомолога на другой хромосоме. Обе хромосомы <