Передача возбуждения с нервного волокна на мышечное осуществляется через посредство нервно-мышечного синапса (морфо-функциональный контакт между пресинаптическим окончанием аксона эфферентного нейрона и определенным участком мембраны мышечного волокна). По способу передачи информации мионевральный синапс относится к синапсам химического типа.
В нервно-мышечном синапсе, как и в любом другом, выделяют:
Ø пресинаптическую мембрану (часть мембраны пресинаптического окончания аксона двигательного нейрона, вступающая в контакт с мембраной мышечного волокна),
Ø постсинаптическую мембрану (часть мембраны мышечного волокна, с которой вступает в контакт мембрана нервного волокна),
Ø синаптическую щель (узкое пространство между пре- и постсинаптической мембранами, заполненное межклеточной жидкостью; ширина этого пространства в нервно-мышечном синапсе составляет 80-100 нм).
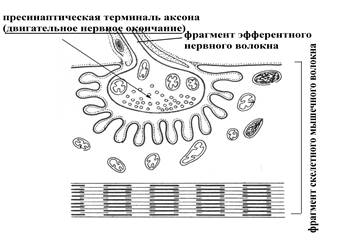

А Б
Рис. 8. Мотонейрон (двигательный нейрон) и иннервируемое им скелетное мышечное волокно (А); прямоугольник указывает на нервно-мышечный синапс (Б – его схематическое представление), в образовании которого принимает участие пресинаптическая терминаль аксона мотонейрона (по Е.А. Шубниковой, 1987).
Нервно-мышечный синапс является химическим: передача возбуждения с нервного волокна на мышечное (будь-то скелетное или гладкое мышечное волокно) осуществляется с участием специальных химических посредников – медиаторов (химических веществ, чаще всего, органической природы). Так, медиатором в синапсе между нервным и скелетным мышечным волокнами является ацетилхолин (Ах), а в синапсах между нервными и гладкими мышечными волокнами – могут выступать как ацетилхолин, так и норадреналин (НА).
Рассмотрим механизм передачи возбуждения с нервного волокна на скелетное мышечное волокно (этот механизм является в определенном плане универсальным для всех химических синапсов, но в зависимости от того, какие структуры принимают участие в образовании синапса, имеет свои особенности).
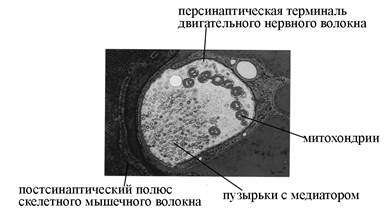

Б
А
В

Рис. 6. Ультраструктурная организация нервно-мышечного синапса (по T. Smit, 2002). А – СЭМ терминали двигательного нервного волокна и фрагмента скелетного мышечного волокна, принимающего участие в образовании синаптического контакта. Б – ТЭМ нервно-мышечного синапса. В – схема строения химического синапса.
Возбуждение в виде нервного импульса распространяется от тела нервной клетки вдоль ее аксона к пресинаптическому окончанию аксона. При этом на пресинаптической мембрана аксона возникает не потенциал действия, а локальная деполяризация. В пресинаптической мембране, наряду с электровозбудимыми Na+- и К+-каналами, имеются и электровозбудимые (потенциалзависимые) Са2+-каналы, срабатывающие на деполяризацию открытием своих единственных ворот. Как следствие этого, проницаемость пресинаптической мембраны для ионов кальция повышается, и они начинают входить по концентрационному градиенту из синаптической щели в пресинаптическое окончание аксона, в результате чего концентрация кальция внутри пресинаптического окончания возрастает. В пресинаптическом окончании аксона имеются актиновые филаменты, которые ассоциированы с мембранными пузырьками, содержащими медиатор (ацетилхолин), и миозиновые филаменты, которые одним своим концом прикреплены к внутренней поверхности пресинаптической мембраны, а другой их конец свободен и направлен в сторону пузырьков с медиатором. Повышение концентрации кальция в пресинаптическом окончании, в свою очередь, делает возможным взаимодействие актиновых и миозиновых филаментов между собой, что сопровождается продвижением пузырьков с медиатором в направлении внутренней поверхности пресинаптической мембраны.

А Б
Рис. 7. Схематическое изображение хемовозбудимого канала. А – закрытое состояние, в отсутствии связи ацетилхолина (Ах) с рецептором, Б – открытое состояние канала, возникающее после присоединения ацетилхолина к холинорецептору.
В момент, когда пузырьки, содержащие ацетилхолин, приближаются к пресинаптической мембране на определенное расстояние, их мембрана встраивается в пресинаптическую, а содержимое (медиатор – ацетилхолин) оказывается в синаптической щели. Количество молекул ацетилхолина во всех пузырьках примерно одинаково; содержимое одного пузырька получило название кванта медиатора. Таким образом, деполяризация пресинаптической мембраны сопровождается выделением в синаптическую щель определенного количества квантов медиатора ацетилхолина.
Ацетилхолин диффундирует через синаптическую щель и достигает постсинаптической мембраны. Особенностью постсинаптической мембраны является отсутствие потенциалзависимых каналов (имеющихся во всей остальной мембране мышечного волокна, называемой внесинаптической), но наличие хемовозбудимых каналов. Состояние хемовозбудимых каналов не зависит от величины мембранного потенциала, но может изменяться под влиянием определенных химических веществ (в частности, молекул медиатора).
В хемовозбудимом канале, подобно электровозбудимому, различают отверстие (пору), в области которого расположены ворота (белковой природы); но состояние этих ворот (их пространственная структура, определяющая открытое или закрытое состояние канала) зависит не от величины заряда на мембране, а от состояния, расположенного вблизи этих ворот белка-рецептора, обладающего высоким сродством к медиатору (и возможно некоторым другим веществам). В частности, в хемовозбудимых каналах постсинаптической мембраны скелетного мышечного волокна имеются белковые холинорецептор и ворота. В отсутствии ацетилхолина в области холинорецептора канал закрыт. Присоединение ацетилхолина к холинорецептору сопровождается изменением его конформации, что приводит и к изменению конформации расположенных поблизости от холинорецептора ворот таким образом, что они отодвигаются от поры канала, и канал открывается. Размер пор в хемовозбудимых каналах (~0,65 нм) несколько больше такового в электровозбудимых натриевых и калиевых каналах, в связи с чем они потенциально способны пропускать любые ионы (натрия, калия, кальция), т.е. характеризуются сравнительно низкой селективностью. Вместе с тем преобладающими в силу распределения электрического заряда на мембране является входящий натриевый ток, который деполяризует постсинаптическую мембрану мышечного волокна (возникает т.н. постсинаптический потенциал). Но такая деполяризация не может приобретать самообновляющийся характер, поскольку в постсинаптической мембране отсутствуют потенциалзависимые каналы. Доля открытых хемовозбудимых каналов определяется количеством высвободившихся в синаптическую щель молекул медиатора. В связи с этим постсинаптический потенциал, в отличие от потенциала действия, является градуальным. При некоторой определенной амплитуде постсинаптического потенциала в ближайшем к синапсу участке внесинаптической мембраны мышечного волокна активная деполяризация, вызванная этим постсинаптическим потенциалом, приобретает самообновляющийся характер, что приводит к генерации нервного импульса, который проводится вдоль скелетного мышечного волокна, инициируя его сокращение.
Важную роль в функционировании нервно-мышечного синапса играет фермент холинэстераза, который локализован в синаптической щели и расщепляет ацетилхолин. Расщепление ацетилхолина обеспечивает не постоянное, а временное его действие на постсинаптическую мембрану, благодаря чему синаптическая передача носит дискретный характер. При ритмическом слишком частом следовании нервных импульсов к пресинаптическому окончанию по нервному волокну холинэстераза не успевает расщепить ацетилхолин, выделившейся при каждом предыдущем возбуждении пресинаптической мембраны нервного волокна, и все бóльшая доля хемовозбудимых каналов постсинаптической мембраны скелетного мышечного волокна оказывается открытой, что приводит к росту постсинаптического потенциала. Данное обстоятельство является причиной длительной и стойкой деполяризации внесинаптической мембраны мышечного волокна, сопровождающейся постепенной натриевой инактивацией и снижением возбудимости внесинаптической мембраны вплоть до полной временной утраты способности генерировать нервные импульсы; наступает блок синаптической передачи (пессимальное торможение нервно-мышечного синапса). Пессимальное торможение нервно-мышечной передачи наступает при частоте стимуляции нервного волокна 100-150 имп./с. Таким образом, лабильность нервно-мышечного синапса в связи с химическим механизмом передачи гораздо меньше таковой нервного волокна (до 2000 имп./с) и скелетных мышечных волокон (до 500 имп./c).
Химический механизм передачи возбуждения через синапс определяет следующие ее особенности:
Ø одностороннее проведение возбуждения (только в направлении от нервного волокна на мышечное волокно). Связано с тем, что синаптическая щель в химических синапсах довольно широкая (до 80-100 нм), и ионные токи, возникающие при деполяризации постсинаптической мембраны, шунтируются в этой щели и не способны вызвать возвратную деполяризацию пресинаптической мембраны
Ø задержанное проведение возбуждения (латентный период передачи информации через синапс составляет 0,3-0,5 мс). Обусловлено тем, что для осуществления процессов выделения медиатора в синаптическую щель под действием приходящего к пресинапсу нервного импульса, диффузии медиатора к постсинаптической мембране и проявления его эффектов на постсинаптическую мембрану требуется определенное время
Ø относительно низкая лабильность синапсов (по сравнению с нервными и мышечными волокнами) в связи с зависимостью процесса нервно-мышечной передачи от медиатора и необходимостью для нормальной работы синапса постоянного расщепления медиатора после каждого проводимого возбуждения
Ø относительно высокая утомляемость синапсов (поскольку максимальная продолжительность их постоянного функционирования лимитируется запасом "готового" к высвобождению медиатора; кроме того, при длительном проведении возбуждения через синапс уменьшается чувствительность постсинаптической мембраны к ацетилхолину, что также является одной из причин развития утомления в синапсе).
Химический способ передачи информации через нервно-мышечный синапс обуславливает, наряду с отмеченным, и высокую степень зависимости этого процесса от ряда химических веществ (т.н. модуляторов синаптической передачи). Так, временно заблокировать нервно-мышечную передачу можно несколькими путями:
Ø блокируя секрецию ацетилхолина в синаптическую щель (таким действием обладают столбнячный и ботулинический токсины),
Ø блокируя холинорецепторы постсинаптической мембраны (таким действием обладают кураре и его производные),
Ø инактивируя холинэстеразу (например, под действием прозерина).