Под нервным центром понимают сложный комплекс (ансамбль) нейронов, совокупно участвующих в регуляции какой-то физиологической функции или осуществлении определенного рефлекторного акта. Существует "понимание" нервных центров в узком и широком смысле этого понятия. Под нервными центрами в узком смысле понимают лишь те структуры центральной нервной системы, которые являются жизненно необходимыми для осуществления или регуляции определенного физиологического процесса, тогда как нервный центр в широком смысле этого понятия представляет собой совокупность всех компонентов центральной нервной системы, имеющих отношение к осуществлению и регуляции определенного физиологического процесса. Так, например, жизненно важный дыхательный центр, обеспечивающий ритмическую смену вдоха выдохом, находится в продолговатом мозге, но в то же время сами мотонейроны, иннервирующие дыхательную мускулатуру, заложены в спинном мозге. Кроме того, нервные клетки, имеющие отношение к регуляции внешнего дыхания, находятся в мосте, гипоталамусе (части промежуточного мозга) и коре больших полушарий. В связи с отмеченным под дыхательным центром в узком смысле этого слова понимают лишь структуры продолговатого мозга, имеющие отношение к периодической смене вдоха выдохом, тогда как в широком смысле этого понятия – совокупность всех нервных элементов центральной нервной системы, имеющих отношение к регуляции дыхания.
Для всех нервных центров характерны следующие свойства:
1. Одностороннее проведение возбуждения (возбуждение проводится всегда в одном направлении от афферентного нейрона к эфферентному, а затем к структурам исполнительных органов). Достигается благодаря существованию синапсов химического типа между чувствительными, вставочными и эфферентными нейронами.
2. Замедленное проведение возбуждения через нервные центры. Обусловлено передачей информации от одних нейронов центральной нервной системы к другим, имеющим отношение к регуляции какой-то физиологической функции, преимущественно через химические синапсы, для которых характерна синаптическая задержка (0,3-0,5 мс при проведении информации через один синапс). Соответственно, чем больше синаптических контактов встречается на пути нервного импульса от чувствительного нейрона к эфферентному, тем длительнее будет общая задержка проведения через соответствующий нервный центр.
3. Зависимость рефлекторного ответа от силы и длительности раздражения рецептивного поля рефлекса. Существование такой связи обусловлено тем, что, чем сильнее действующий на рецептивное поле какого-то рефлекторного акта раздражитель и чем больше длительность его действия, тем соответственно больше будет количество элементов нервного центра, вовлекаемых в реакцию, и большим окажется количество структур иннервируемых рабочих органов, участвующих в рефлекторном ответе, а, следовательно, и выше будет амплитуда рефлекторного ответа.
4. Суммация возбуждений в нервном центре. Это свойство проявляется следующим образом: раздражение каких-то отдельных рецепторов рецептивного поля нервного центра слабыми раздражителями не вызывает возникновения рефлекторного ответа, тогда как последовательное с определенной частотой раздражение этого же рецептора или одновременное воздействие на несколько соседних рецепторных элементов теми же слабыми стимулами, приводит к возникновению рефлекторного ответа. В основе описанного феномена лежат явления суммации возбуждений, поступающих к одному и тому же центральному нейрону либо через какой-то один из его синаптических входов с определенной частотой (временная суммация), либо одновременно через несколько его синаптических входов (пространственная суммация).
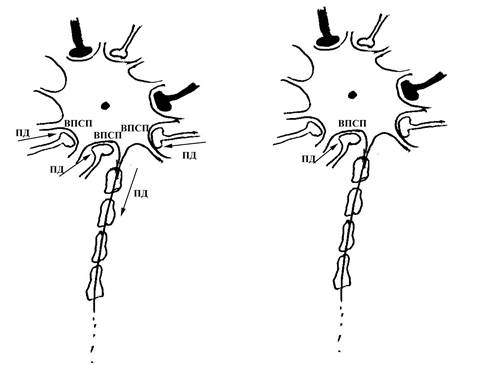
А Б
Рис. 68. Схема пространственной суммации информации, поступающей к мембране тела нейрона по различным его синаптическим входам. Обратите внимание на то, что на рисунке схематически изображен нейрон, на теле которого расположено большое количество синапсов (незатушеванные – возбуждающие, затушеванные – тормозные). На рисунке А показан такой вариант, когда к пресинаптическим полюсам сразу нескольких синапсов (в данном случае трех возбуждающих синапсов) одновременно приходит возбуждение, которое инициирует возникновение в постсинаптической мембране этих синапсов (являющейся частью мембраны тела рассматриваемого нейрона) возбуждающих постсинаптических потенциалов (ВПСП), суммирующихся в пространстве. Суммарный возбуждающий постсинаптический потенциал, возникающий в результате такой суммации, является достаточным для инициации потенциала действия (ПД) в области аксонного холмика данного нейрона.
На рисунке Б показан вариант, когда возбуждение приходит только к одному из пресинаптических полюсов возбуждающих синапсов, поэтому ВПСП возникает только в области постсинаптической мембраны данного синапса, и сам по себе оказывается недостаточным для инициации ПД в области аксонного холмика нейрона.
5. Трансформация ритма возбуждений. Заключается в том, что количество нервных импульсов, поступающих к какому-либо центральному нейрону (например, по чувствительным нервным волокнам), как правило, всегда отлично от такового, возникающего в ответ в этом нейроне. Так, частота импульсации, проводимой по чувствительным нервным волокнам, как правило, отлична от той, которая возникает во вставочных и эфферентных нервных волокнах. Отмеченное обстоятельство обусловлено разной лабильностью и электрофизиологическими свойствами (например, неодинаковой продолжительностью следовых потенциалов, сопровождающих пик потенциала действия) различных нервных элементов нервного центра
6. Рефлекторное последействие. Проявляется в том, что собственно рефлекторный ответ на действие раздражителя прекращается не мгновенно после прекращения раздражения соответствующего рецептивного поля рефлекса, а спустя какое-то время, что свидетельствует в пользу некоторой инерции возбуждения в нервном центре. Такая инерция возбуждения в нервном центре может быть следствием, как минимум, двух обстоятельств. Во-первых, возбуждение вставочного или эфферентного нейрона, возникающее в результате передачи информации от чувствительного нейрона, не исчезает сразу же после прекращения возбуждения чувствительного нейрона, а может сохраняться некоторое время благодаря выраженной следовой деполяризации в нейронах нервного центра после каждого потенциала действия, вызывающей возникновение новых импульсных ответов. Во-вторых, инерция возбуждения в нервных центрах может быть обусловлена способностью возбуждения циркулировать по замкнутым нейронным цепям нервного центра.
 |
Рис. 69. Схема циркуляции возбуждения по замкнутым нервным цепям в нервных центрах (по Бабскому Е.Б, 1972).
7. Утомление нервных центров. Состоит в снижении работоспособности нервных центров после длительного периода постоянной их активности и, как следствие, временном ослаблении или полном прекращении регуляции какой-то физиологической функции. Более высокая утомляемость нервных центров, по сравнению с нервными клетками, обусловлена наличием синаптических контактов химического типа между различными нейронными элементами в пределах нервного центра.
8. Тонус нервных центров, проявляющийся в постоянном их возбуждении. Возникает и поддерживается в большинстве нервных центров по причине постоянного поступления информации по афферентным нейронам от исполнительных органов рефлекторных актов (т.е. наличию постоянной афферентации от исполнительных органов в нервный центр).
9. Избирательная повышенная чувствительность нервных центров к определенным веществам, нарушающим их нормальную работу (т.н. нервным ядам). Предполагают, что нервные яды нарушают нормальное протекание обмена веществ в нервных центрах, действуя на определенные ферментативные системы их нейронов. Так, например, рвотный центр обладает высокой чувствительностью к апоморфину: это вещество вызывает стойкое возбуждение рвотного центра и связанную с этим длительную рвоту.
10. Существование постоянной обратной связи между нервным центром и исполнительными органами. Достигается благодаря постоянному поступлению по чувствительным нервным волокнам к нервному центру информации о функциональном состоянии рабочих (исполнительных) их органов, что, с одной стороны, обеспечивает поддержание постоянного тонуса (степени возбуждения нервного центра), а с другой – очень тонкую регуляцию деятельности исполнительных органов, поскольку эфферентные нервные посылки к органам постоянно модифицируются в зависимости от изменения функционального состояния исполнительных органов в момент осуществления определенного рефлекторного акта и соответственно поступающей к нервным центрам афферентной информации о состоянии регулируемых органов.
11. Пластичность нервных центров, проявляющаяся в компенсаторной их приспособляемости к повреждению и возможности функциональной перестройки. Так, частичное повреждение нервного центра вначале сопровождается возникновением определенных расстройств в регулируемой физиологической функции, которые со временем полностью или частично проходят по причине компенсаторных изменений в оставшихся нормальными структурах нервного центра, восполняющих функцию поврежденных структур. Функциональная перестройка нервных центров возникает также в случае искусственного, вызванного хирургическим вмешательством, изменения связи нервных центров с исполнительными органами. Так, например, перерезка каких-то нервов, иннервирующих различные органы, и последующее перекрестное их сшивание, в результате чего нервы начинают иннервировать другие внутренние органы, со временем сопровождается тем, что нервные центры начинают нормально регулировать деятельность других органов, с которыми оказались связанными (т.е. нервные центры приобретают новые, ранее им не свойственные, функции). Так, например, при параличе лицевого нерва иногда производят операцию по сшиванию периферического конца этого нерва (собственно вступающего в контакт с мимической мускулатурой лица) с какими-либо другими нормальными черепно-мозговыми нервами, и через некоторое время функция неработающих по причине паралича лицевого нерва мимических мышц лица восстанавливается.