Строение и проводящие пути спинного мозга
Строение спинного мозга
Спинной мозг, medulla spinalis (греч. myelos), лежит в позвоночном канале и у взрослых представляет собой длинный (45 см у мужчин и 41—42 см у женщин), несколько сплюснутый спереди назад цилиндрический тяж, который вверху (краниально) непосредственно переходит в продолговатый мозг, а внизу (каудально) оканчивается коническим заострением, conus medullaris, на уровне II поясничного позвонка.
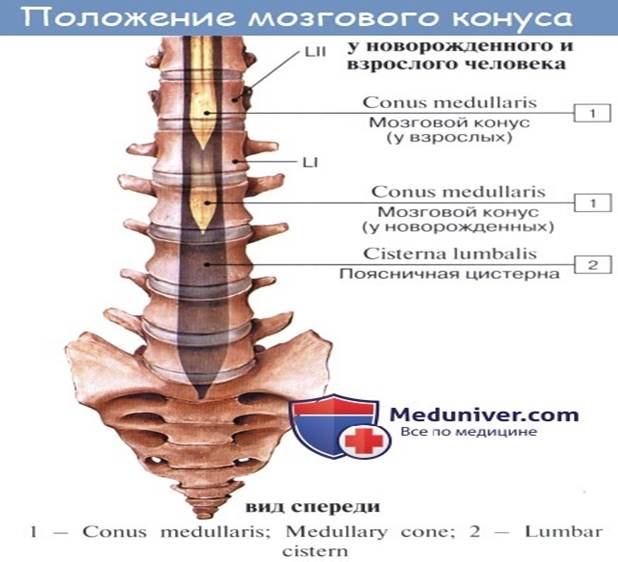
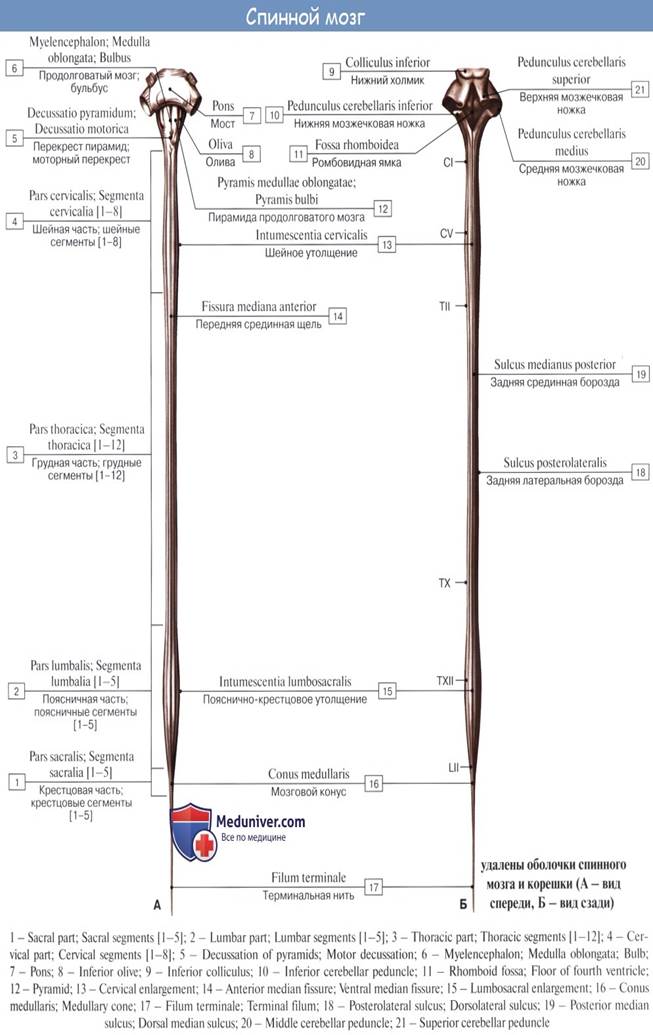
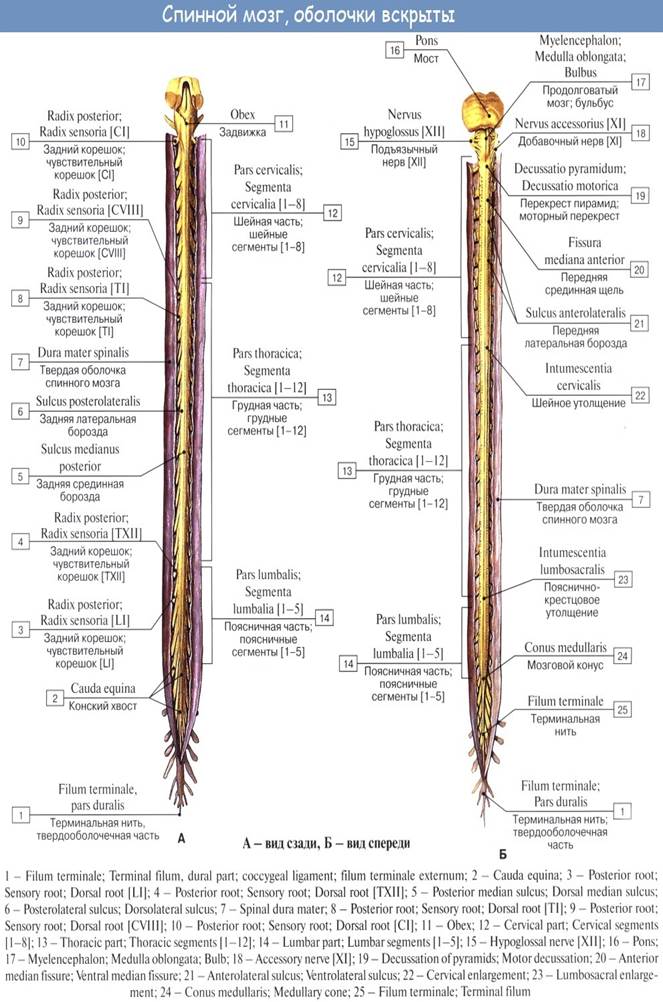
Знание этого факта имеет практическое значение (чтобы не повредить спинной мозг при поясничном проколе с целью взятия спинномозговой жидкости или с целью спинномозговой анестезии, надо вводить иглу шприца между остистыми отростками III и IV поясничных позвонков). От conus medullaris отходит книзу так называемая концевая нить, filum terminale, представляющая атрофированную нижнюю часть спинного мозга, которая внизу состоит из продолжения оболочек спинного мозга и прикрепляется ко II копчиковому позвонку.
Спинной мозг на своем протяжении имеет два утолщения, соответствующих корешкам нервов верхней и нижней конечностей: верхнее из них называется шейным утолщением, intumescentia cervicalis, а нижнее — пояснично-крестцовым, intumescentia lumbosacralis.
Из этих утолщений более обширно пояснично-крестцовое, но более дифференцировано шейное, что связано с более сложной иннервацией руки как органа труда. Образовавшимися вследствие утолщения боковых стенок спинномозговой трубки и проходящими по средней линии передней и задней продольными бороздами:
глубокой fissura mediana anterior,
и поверхностной, sulcus medianus posterior,
спинной мозг делится на две симметричные половины — правую и левую; каждая из них в свою очередь имеет слабо выраженную продольную борозду, идущую по линии входа задних корешков (sulcus posterolateralis) и по линии выхода передних корешков (sulcus anterolateralis).
Проводящие пути спинного мозга
В нервной системе нервные клетки не лежат изолированно. Они вступают в контакт друг с другом, образуя цепи нейронов - проводников импульсов. Длинный отросток одного нейрона - нейрит (аксон) вступает в контакт с короткими отростками (дендритами) или телом другого, следующего в цепи нейрона.
По цепям нейронов нервные импульсы движутся в строго определенном направлении, что обусловлено особенностями строения нервных клеток и синапсов («динамическая поляризация»). Одни цепи нейронов несут импульс в центростремительном направлении - от места возникновения на периферии (в коже, слизистых оболочках, органах, стенках сосудов) к ЦНС (спинному и головному мозгу). Первым в этой цепи является чувствительный (афферентный) нейрон, воспринимающий раздражение и трансформирующий его в нервный импульс. Другие цепи нейронов проводят импульс в центробежном направлении - от головного или спинного мозга на периферию, к рабочему органу. Нейрон, передающий импульс рабочему органу, является эфферентным.
Цепи нейронов в живом организме образуют рефлекторные дуги.
Рефлекторная дуга - это цепь нервных клеток, обязательно включающая первый - чувствительный и последний - двигательный (или секреторный) нейроны, по которым импульс движется от места возникновения к месту приложения (мышцы, железы и другие органы, ткани).
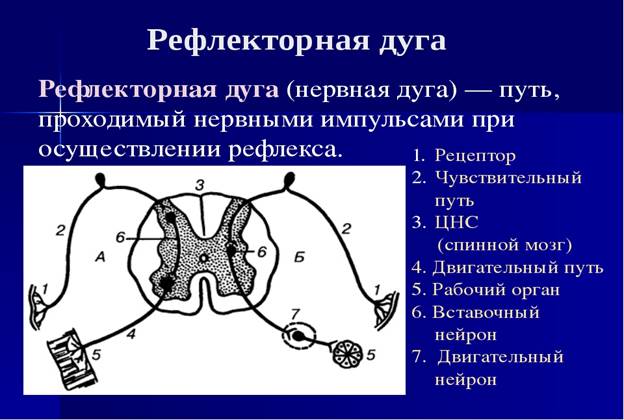
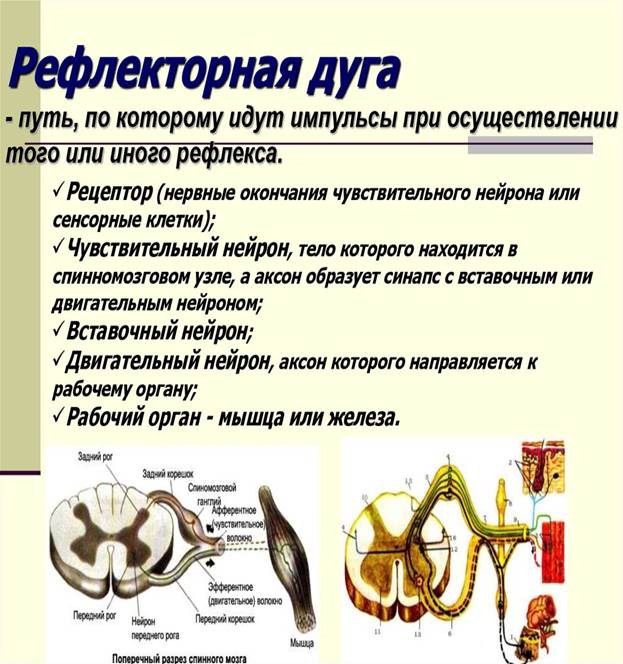
Наиболее простыми рефлекторными дугами являются двух- и трехнейронные, замыкающиеся на уровне одного сегмента спинного мозга. В трехнейронной рефлекторной дуге первый нейрон представлен чувствительной клеткой, по которой импульс от места возникновения в чувствительном нервном окончании (рецепторе), лежащем в коже или в других органах, движется вначале по периферическому отростку (в составе нерва). Затем импульс движется по центральному отростку в составе заднего корешка спинномозгового нерва, направляясь к одному из ядер заднего рога спинного мозга, или по чувствительным волокнам черепных нервов к соответствующим чувствительным ядрам.
Здесь импульс передается следующему нейрону, отросток которого направляется из заднего рога в передний, к клеткам ядер (двигательных) переднего рога. Этот второй нейрон выполняет проводниковую (кондукторную) функцию. Он передает импульс от чувствительного (афферентного) нейрона к третьему - двигательному (эфферентному). Кондукторный нейрон является вставочным нейроном, так как находится между чувствительным нейроном, с одной стороны, и двигательным (или секреторным) - с другой. Тело третьего нейрона (эфферентного, эффекторного, двигательного) лежит в переднем роге спинного мозга, а его аксон - в составе переднего корешка, а затем спинномозгового нерва простирается до рабочего органа (мышцы).
С развитием спинного и головного мозга усложнились и связи в нервной системе. Образовались многонейронные сложные рефлекторные дуги, в построении и функциях которых участвуют нервные клетки, расположенные в вышележащих сегментах спинного мозга, в ядрах мозгового ствола, полушарий и даже в коре большого мозга. Отростки нервных клеток, проводящих нервные импульсы из спинного мозга к ядрам и коре головного мозга и в обратном направлении, образуют пучки (fasciculi).
Пучки нервных волокон, соединяющие функционально однородные или различные участки серого вещества в ЦНС, занимающие в белом веществе головного и спинного мозга определенное место и проводящие одинаковый импульс, получили название проводящих путей.
В спинном и головном мозге по строению и функции выделяют три группы проводящих путей:
1. ассоциативные,
2. комиссуральные,
3. проекционные.
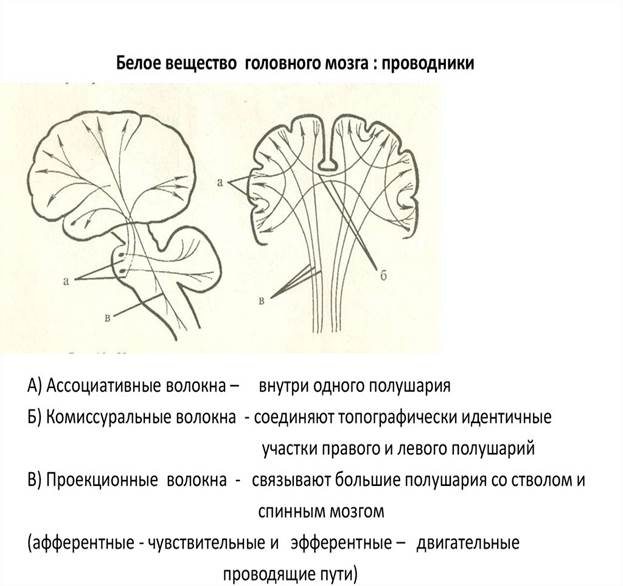
v Ассоциативные нервные волокна (neurofibrae associations) соединяют участки серого вещества, различные функциональные центры (кора мозга, ядра) в пределах одной половины мозга.
Выделяют короткие и длинные ассоциативные волокна (пути). Короткие волокна соединяют близлежащие участки серого вещества и располагаются в пределах одной доли мозга (внутридолевые пучки волокон).
Некоторые ассоциативные волокна, соединяющие серое вещество соседних извилин, не выходят за пределы коры (интракортикальные). Они дугообразно изгибаются в виде буквы 0 и называются дугообразными волокнами большого мозга (fibrae arcuatae cerebri). Ассоциативные нервные волокна, выходящие в белое вещество полушария (за пределы коры), называют экстракортикальными.
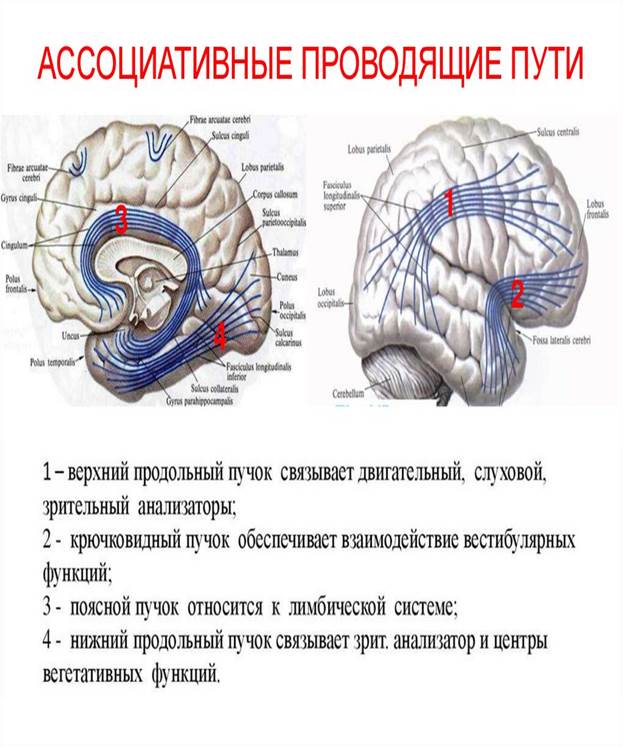
Длинные ассоциативные волокна связывают участки серого вещества, далеко отстоящие друг от друга, принадлежащие различным долям (междолевые пучки волокон). Это хорошо выраженные пучки волокон, которые можно видеть на макропрепарате головного мозга. К длинным ассоциативным путям относятся следующие: верхний продольный пучок (fasciculus longitudinalis superior), который находится в верхней части белого вещества полушария большого мозга и соединяет кору лобной доли с теменной и затылочной; нижний продольный пучок (fasciculus longitudinalis inferior), лежащий в нижних отделах полушария и соединяющий кору височной доли с затылочной; крючков,идный пучок (fasciculus uncinatus), который, дугообразно изгибаясь впереди островка, соединяет кору в области лобного полюса с передней частью височной доли. В спинном мозге ассоциативные волокна соединяют клетки серого вещества, принадлежащего различным сегментам, и образуют передние, латеральные и задние собственные пучки (межсегментные пучки) (fasciculi proprii ventrales, s. anteriores lateralis, dorsrales, s. posteriores). Они располагаются непосредственно возле серого вещества. Короткие пучки связывают соседние сегменты, перекидываясь через 2-3 сегмента, длинные пучки соединяют далеко отстояшие друг от друга сегменты спинного мозга.
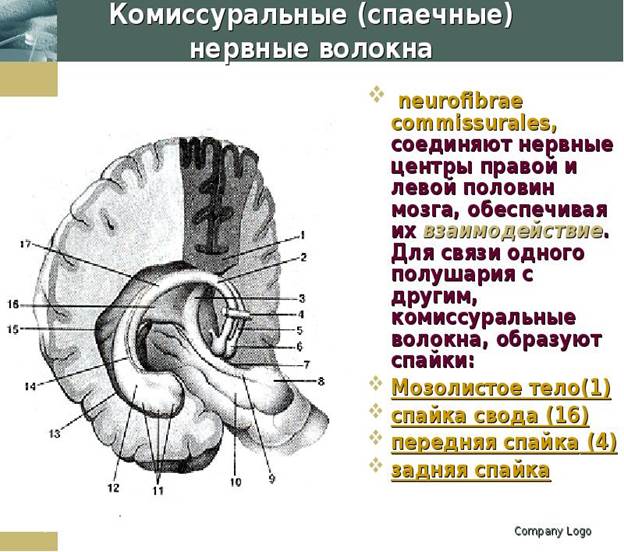
v Комиссуральные (спаечные) нервные волокна (neurofibrae commissurales) соединяют серое вещество правого и левого полушарий, аналогичные центры правой и левой половин мозга с целью координации их функций.
Комиссуральные волокна проходят из одного полушария в другое, образуя спайки (мозолистое тело, спайка свода, передняя спайка). В мозолистом теле, имеющемся только у млекопитающих, располагаются волокна, соединяющие новые, более молодые, отделы мозга, корковые центры правого и левого полушарий.
В белом веществе полушарий волокна мозолистого тела расходятся веерообразно, образуя лучистость мозолистого тела (radiatio corporis callosi).
Комиссуральные волокна, идущие в колене и клюве мозолистого тела, соединяют друг с другом участки лобных долей правого и левого полушарий большого мозга. Загибаясь кпереди, пучки этих волокон как бы охватывают с двух сторон переднюю часть продольной щели большого мозга и образуют лобные щипцы (forceps frontalis). В стволе мозолистого тела проходят нервные волокна, соединяющие кору центральных извилин, теменных и височных долей двух полушарий большого мозга. Валик мозолистого тела состоит из комиссуральных волокон, которые соединяют кору затылочных и задние отделы теменных долей правого и левого полушарий большого мозга. Изгибаясь кзади, пучки этих волокон охватывают задние отделы продольной щели большого мозга и образуют затылочные щипцы (forceps occipitalis).
Комиссуральные волокна проходят в составе передней спайки мозга (commissura rostralis, s. anterior) и спайки свода (commissura fornicis).
Большая часть комиссуральных волокон, входящих в состав передней спайки, - это пучки, соединяющие друг с другом переднемедиальные участки коры височных долей обоих полушарий в дополнение к волокнам мозолистого тела.
В составе передней спайки находятся также слабовыраженные у человека пучки комиссуральных волокон, направляющиеся из области обонятельного треугольника одной стороны мозга в такую же область другой стороны.
В спайке свода проходят комиссуральные волокна, которые соединяют участки коры правой и левой височных долей полушарий большого мозга, правого и левого гиппокампов.
v Проекционные нервные волокна (neurofibrae proectiones) соединяют нижележащие отделы мозга (спинной мозг) с головным мозгом, а также ядра мозгового ствола с базальными ядрами (полосатым телом) и корой и, наоборот, кору головного мозга, базальные ядра с ядрами мозгового ствола и со спинным мозгом.
При помощи проекционных волокон, достигающих коры большого мозга, картины внешнего мира как бы проецируются на кору как на экран, где происходят высший анализ поступивших сюда импульсов, сознательная их оценка.
В группе проекционных путей выделяют восходящие и нисходящие системы волокон.
Восходящие проекционные пути (афферентные, чувствительные) несут в головной мозг, к его подкорковым и высшим центрам (к коре), импульсы, возникшие в результате воздействия на организм факторов внешней среды, в том числе и от органов чувств, а также импульсы от органов движения, внутренних органов, сосудов. По характеру проводимых импульсов восходящие проекционные пути подразделяются на три группы.
Экстероцептивные пути (от лат. exter. externus - наружный, внешний) несут импульсы (болевые, температурные, осязания и давления), возникшие в результате воздействия внешней среды на кожные покровы, а также импульсы от высших органов чувств (органов зрения, слуха, вкуса, обоняния).
Проприоцептивные пути (от лат. proprius - собственный) проводят импульсы от органов движения (от мышц, сухожилий, суставных капсул, связок), несут информацию о положении частей тела, о размахе движений.
Интероцептивные пути (от лат. interior - внутренний) проводят импульсы от внутренних органов, сосудов, где хемо-, баро- и механорецепторы воспринимают состояние внутренней среды организма, интенсивность обмена веществ, химизм крови, тканевой жидкости, лимфы, давление в сосудах.
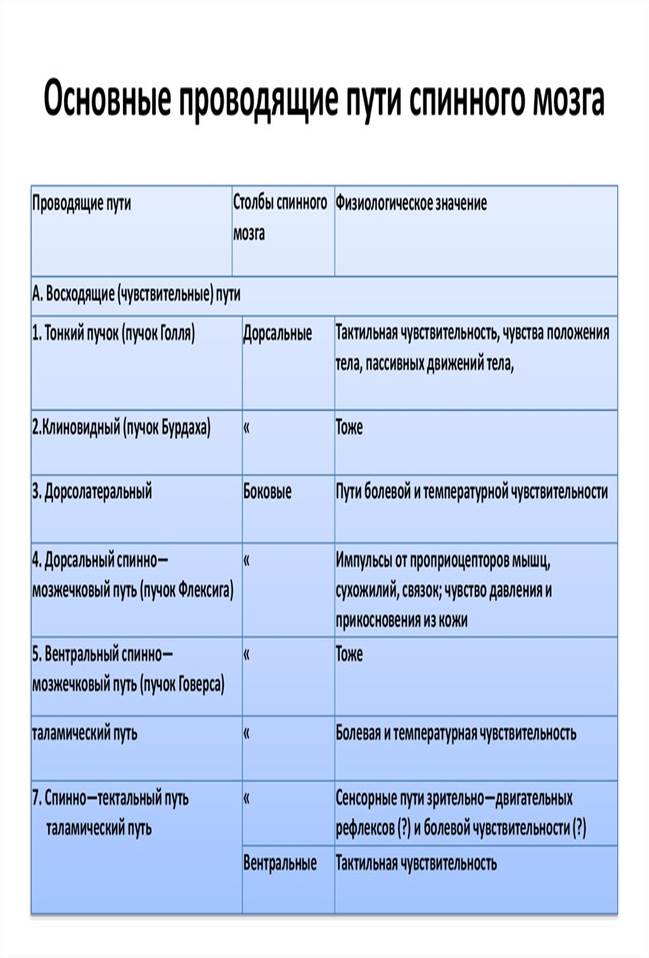
Экстероцептивные проводящие пути. Проводящий путь болевой и температурной чувствительности - латеральный спинно-таламический путь (tractus spinothalamicus lateralis) состоит из трех нейронов.
Чувствительным проводящим путям принято давать названия с учетом топографии - места начала и конца второго нейрона. Например, у спинно-таламического пути второй нейрон простирается от спинного мозга, где в заднем роге лежит тело клетки, до таламуса, где аксон этого нейрона образует синапс с клеткой третьего нейрона.
Рецепторы первого (чувствительного) нейрона, воспринимающие чувство боли, температуру, располагаются в коже, слизистых оболочках, а нейрит третьего нейрона заканчивается в коре постцентральной извилины, где находится корковый конец анализатора общей чувствительности.
Тело первой чувствительной клетки лежит в спинномозговом узле, а ее центральный отросток в составе заднего корешка направляется в задний рог спинного мозга и заканчивается синапсами на клетках второго нейрона.
Аксон второго нейрона, тело которого лежит в заднем роге, направляется на противоположную сторону спинного мозга через его переднюю серую спайку и входит в боковой канатик, где включается в состав латерального спинно-таламического пути.
Из спинного мозга пучок поднимается в продолговатый мозг и располагается позади ядра оливы, а в покрышке моста и среднего мозга лежит у наружного края медиальной петли.
Заканчивается второй нейрон латерального спинно-таламического пути синапсами на клетках дорсального латерального ядра таламуса.
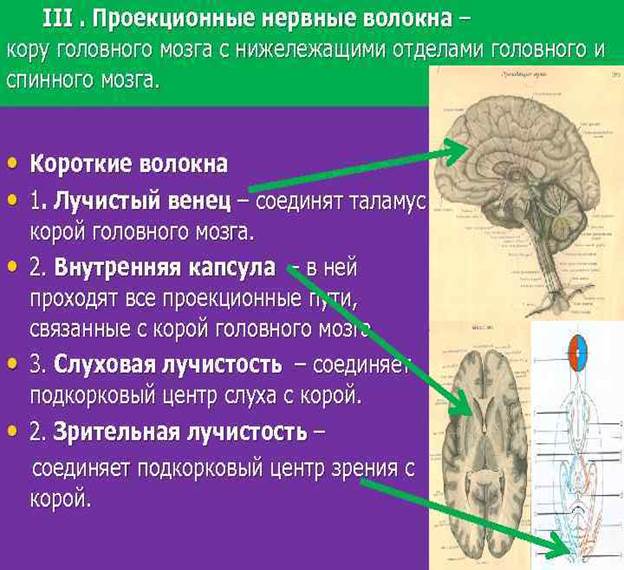
Здесь расположены тела третьего нейрона, отростки клеток которого проходят через заднюю ножку внутренней капсулы и в составе веерообразно расходящихся пучков волокон, образующих лучистый венец (corona radiata).
Эти волокна достигают коры полушария большого мозга, его постцентральной извилины. Здесь они заканчиваются синапсами с клетками четвертого слоя (внутренняя зернистая пластинка).
Волокна третьего нейрона чувствительного (восходящего) проводящего пути, соединяющего таламус с корой, образуют таламокорковые пучки (fasciculi thalamocorticalis) - таламотеменные волокна (fibrae thalamoparietales).
Латеральный спинно-таламический путь является полностью перекрещенным проводящим путем (все волокна второго нейрона переходят на противоположную сторону), поэтому при повреждении одной половины спинного мозга полностью исчезают болевая и температурная чувствительность на противоположной стороне от повреждения.
Проводящий путь осязания и давления, передний спинно-таламический путь (tractus spinothalamicus ventralis, s. anterior) несет импульсы от кожи, где лежат рецепторы, воспринимающие чувство давления и осязания.
Импульсы идут к коре большого мозга, в постцентральную извилину - место расположения коркового конца анализатора общей чувствительности. Тела клеток первого нейрона лежат в спинномозговом узле, а их центральные отростки в составе заднего корешка спинномозговых нервов направляются в задний рог спинного мозга, где заканчиваются синапсами на клетках второго нейрона.
Аксоны второго нейрона переходят на противоположную сторону спинного мозга (через переднюю серую спайку), входят в передний канатик и в его составе направляются вверх, к головному мозгу.
На своем пути в продолговатом мозге аксоны этого пути присоединяются с латеральной стороны к волокнам медиальной петли и заканчиваются в таламусе, в его дорсальном латеральном ядре, синапсами на клетках третьего нейрона.
Волокна третьего нейрона проходят через внутреннюю капсулу (заднюю ножку) и в составе лучистого венца достигают IV слоя коры постцентральной извилины.
Необходимо отметить, что не все волокна, несущие импульсы осязания и давления, переходят на противоположную сторону в спинном мозге. Часть волокон проводящего пути осязания и давления идет в составе заднего канатика спинного мозга (своей стороны) вместе с аксонами проводящего пути проприоцептивной чувствительности коркового направления. В связи с этим при поражении одной половины спинного мозга кожное чувство осязания и давления на противоположной стороне не исчезает полностью, как болевая чувствительность, а только снижается. Этот переход на противоположную сторону частично осуществляется в продолговатом мозге.
Проприоцептивные проводящие пути. Проводящий путь проприоцептивной чувствительности коркового направления (tractus bulbothalamicus - BNA) называется так, поскольку проводит импульсы мышечно-суставного чувства к коре большого мозга, в постцентральную извилину.
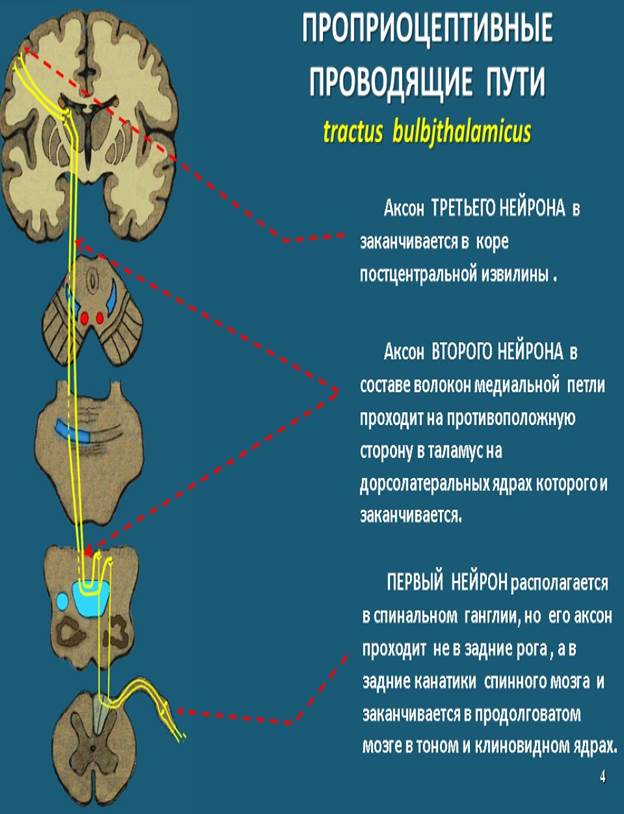
Чувствительные окончания (рецепторы) первого нейрона располагаются в мышцах, сухожилиях, суставных капсулах, связках. Сигналы о тонусе мышц, натяжении сухожилий, о состоянии опорно-двигательного аппарата в целом (импульсы проприоцептивной чувствительности) позволяют человеку оценить положение частей тела (головы, туловища, конечностей) в пространстве, а также во время движения и проводить целенаправленные осознанные движения и их коррекцию. Тела первых нейронов лежат в спинномозговом узле.
Центральные отростки этих клеток в составе заднего корешка направляются в задний канатик, минуя задний рог, а затем уходят вверх в продолговатый мозг к тонкому и клиновидному ядрам.
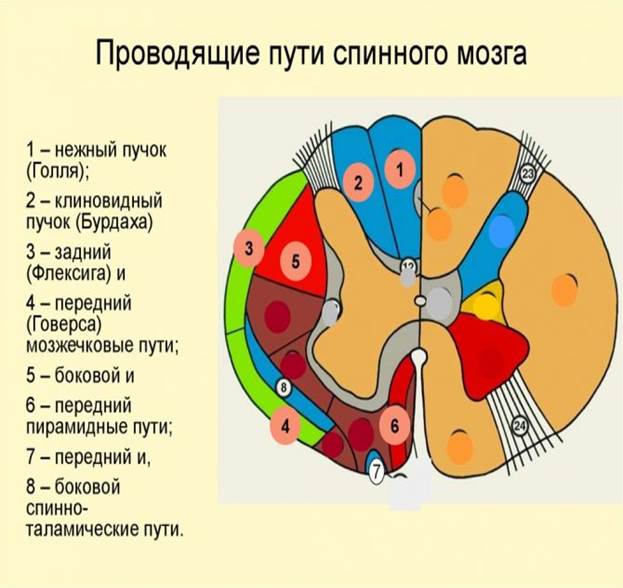
Аксоны, несущие проприоцептивные импульсы, входят в задний канатик начиная с нижних сегментов спинного мозга. Каждый следующий пучок аксонов прилежит с латеральной стороны к уже имеющимся пучкам. Таким образом, наружные отделы заднего канатика (клиновидный пучок, пучок Бурдаха) заняты аксонами клеток, осуществляющих проприоцептивную иннервацию в верхнегрудных, шейных отделах тела и верхних конечностей.
Аксоны, занимающие внутреннюю часть заднего канатика (тонкий пучок, пучок Голля), проводят проприоцептивные импульсы от нижних конечностей и нижней половины туловища.
Центральные отростки первого нейрона заканчиваются синапсами на своей стороне, на клетках второго нейрона, тела которых лежат в тонком и клиновидных ядрах продолговатого мозга.
Аксоны клеток второго нейрона выходят из этих ядер, дугообразно изгибаются вперед и медиально на уровне нижнего угла ромбовидной ямки и в межоливном слое переходят на противоположную сторону, образуя перекрест медиальных петель (decussatio lemniscorum medialis).
Пучок волокон, обращенных в медиальном направлении и переходящих на другую сторону, получил название внутренних дугообразных волокон (fibrae arcuatae internae), которые являются начальным отделом медиальной петли (lemniscus medialis).
Волокна меди альной петли в мосту располагаются в задней его части (в покрышке), почти на границе с передней частью (между пучками волокон трапециевидного тела).
В покрышке среднего мозга пучок волокон медиальной петли занимает место дорсолатеральнее красного ядра, а заканчивается в дорсальном латеральном ядре таламуса синапсами на клетках третьего нейрона.
Аксоны клеток третьего нейрона через заднюю ножку внутренней капсулы и в составе лучистого венца достигают постцентральной извилины.
Часть волокон второго нейрона по выходе из тонкого и клиновидного ядер изгибается кнаружи и разделяется на два пучка. Один пучок - задние наружные дугообразные волокна (fibrae arcuatae externae dorsales, s. posteriores), направляются в нижнюю мозжечковую ножку своей стороны и заканчиваются в коре червя мозжечка. Волокна второго пучка - передние наружные дугообразные волокна (fibrae arcuatae externae ventrales, s. anteriores) уходят вперед, переходят на противоположную сторону, огибают с латеральной стороны оливное ядро и также через нижнюю мозжечковую ножку направляются к коре червя мозжечка. Передние и задние наружные дугообразные волокна несут проприоцептивные импульсы к мозжечку.
Проприоцептивный путь коркового направления также перекрещенный. Аксоны второго нейрона переходят на противоположную сторону не в спинном мозге, а в продолговатом.
При повреждении спинного мозга на стороне возникновения проприоцептивных импульсов (при травме мозгового ствола - на противоположной стороне) теряется представление о состоянии опорно-двигательного аппарата, положении частей тела в пространстве, нарушается координация движений.
Наряду с проприоцептивным проводящим путем, несущим импульсы к коре большого мозга, следует назвать проприоцептивные передний и задний спинно-мозжечковые пути.
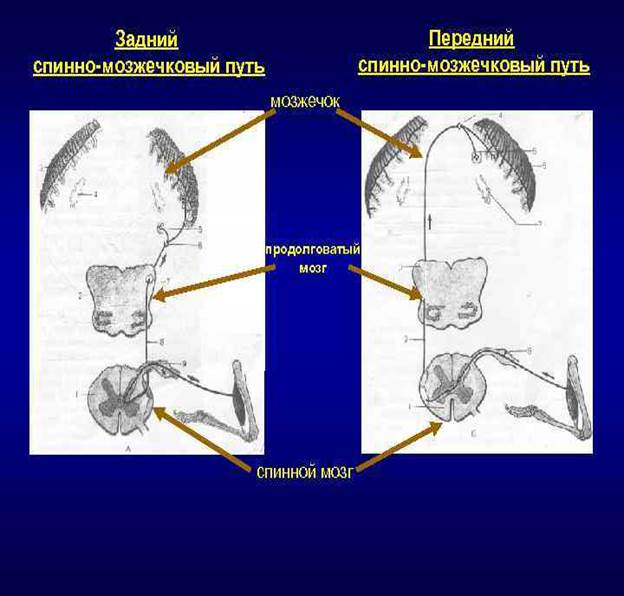
По этим проводящим путям мозжечок получает информацию от расположенных ниже чувствительных центров (спинного мозга) о состоянии опорно-двигательного аппарата, участвует в рефлекторной координации движений, обеспечивающих равновесие тела без участия высших отделов головного мозга (коры полушарий большого мозга).
Задний спинно-мозжечковый путь (tractus spinocerebellaris dorsalis, s. posterior; пучок Флексига) передает проприоцептивные импульсы от мышц, сухожилий, суставов в мозжечок.
Тела клеток первого (чувствительного) нейрона находятся в спинномозговом узле, а центральные отростки их в составе заднего корешка направляются в задний рог спинного мозга и заканчиваются синапсами на клетках грудного ядра (ядра Кларка), лежащего в медиальной части основания заднего рога. Клетки грудного ядра являются вторым нейроном заднего спинно-мозжечкового пути.
Аксоны этих клеток выходят в боковой канатик своей стороны, в его заднюю часть, поднимаются вверх и через нижнюю мозжечковую ножку входят в мозжечок, к клеткам коры червя. Здесь спинно-мозжечковый путь заканчивается.
Можно проследить системы волокон, по которым импульс из коры червя достигает красного ядра, полушария мозжечка и даже вышележащих отделов мозга - коры полушарий большого мозга.
Из коры червя через пробковидное и шаровидное ядра импульс через верхнюю мозжечковую ножку направляется к красному ядру противоположной стороны (мозжечково-покрышечный путь).
Кора червя связана ассоциативными волокнами с корой полушария мозжечка, откуда импульсы поступают в зубчатое ядро мозжечка.
С развитием высших центров чувствительности и произвольных движений в коре полушарий большого мозга возникли также связи мозжечка с корой, осуществляющиеся через таламус.
Таким образом, из зубчатого ядра аксоны его клеток через верхнюю мозжечковую ножку выходят в покрышку моста, переходят на противоположную сторону и направляются к таламусу. Переключившись в таламусе на следующий нейрон, импульс следует в кору большого мозга, в постцентральную извилину.
Передний спинно-мозжечковый путь (tractus spinocerebellaris ventralis, s. anterior; пучок Говерса) имеет более сложное строение, чем задний, поскольку проходит в боковом канатике противоположной стороны, возвращаясь в мозжечок на свою сторону.
Тело клетки первого нейрона располагается в спинномозговом узле. Его периферический отросток имеет окончания (рецепторы) в мышцах, сухожилиях, суставных капсулах. Центральный отросток клетки первого нейрона в составе заднего корешка входит в спинной мозг и заканчивается синапсами на клетках, примыкающих с латеральной стороны к грудному ядру.
Аксоны клеток этого второго нейрона проходят через переднюю серую спайку в боковой канатик противоположной стороны, в его переднюю часть, и поднимаются вверх до уровня перешейка ромбовидного мозга.
В этом месте волокна переднего спинно-мозжечкового пути возвращаются на свою сторону и через верхнюю мозжечковую ножку вступают в кору червя своей стороны, в его передневерхние отделы.
Таким образом, передний спинно-мозжечковый путь, проделав сложный, дважды перекрещенный путь, возвращается на ту же сторону, на которой возникли проприоцептивные импульсы.
Проприоцептивные импульсы, поступившие в кору червя по переднему спинно-мозжечковому проприоцептивному пути, также передаются в красное ядро и через зубчатое ядро в кору большого мозга (в постцентральную извилину).
Нисходящие проекционные пути (эффекторные, эфферентные) проводят импульсы от коры, подкорковых центров к нижележащим отделам, к ядрам мозгового ствола и двигательным ядрам передних рогов спинного мозга. Эти пути можно подразделить на две группы:
1. главный двигательный, или пирамидный путь (корково-ядерный и корково-спинномозговые пути), несет импульсы произвольных движений из коры головного мозга к скелетным мышцам головы, шеи, туловища, конечностей через соответствующие двигательные ядра головного и спинного мозга;
2. экстрапирамидные двигательные пути (tractus rubrospinalis, tractus vestibulospinalis и др.) передают импульсы от подкорковых центров к двигательным ядрам черепных и спинномозговых нервов, а затем к мышцам.

К пирамидному пути (tractus pyramidalis) относится система волокон, по которым двигательные импульсы из коры большого мозга, из предцентральной извилины, от гигантопирамидальных нейронов (клетки Беца) направляются к двигательным ядрам черепных нервов и передним рогам спинного мозга, а от них - к скелетным мышцам.
Учитывая направление хода волокон, а также расположение пучков в стволе головного мозга и канатиках спинного мозга, пирамидный путь подразделяют на три части:
v корково-ядерный - к ядрам черепных нервов;
v латеральный корково-спинномозговой - к ядрам передних рогов спинного мозга;
v передний корково-спинномозговой - также к передним рогам спинного мозга.
Корково-ядерный путь (tractus corticonuclearis) представляет собой пучок отростков гигантопирамидальных нейронов, которые из коры нижней трети предцентральной извилины спускаются к внутренней капсуле и проходят через ее колено. Далее волокна корково-ядерного пути идут в основании ножки мозга, образуя медиальную часть пирамидных путей.
Корково-ядерный, а также корково-спинномозговые пути занимают средние 3/5 основания ножки мозга. Начиная со среднего мозга и далее, в мосту и продолговатом мозге волокна корково-ядерного пути переходят на противоположную сторону к двигательным ядрам черепных нервов: III и IV - в среднем мозге; V, VI, VII - в мосту; IX, X, XI, XII - в продолговатом мозге.
В этих ядрах корково-ядерный путь заканчивается. Составляющие его волокна образуют синапсы с двигательными клетками этих ядер. Отростки упомянутых двигательных клеток выходят из мозга в составе соответствующих черепных нервов и направляются к скелетным мышцам головы и шеи и их иннервируют.
Латеральный и передний корково-спинномозговые пути (tractus corticospinales lateralis et ventralis, s.anterior) также начинаются от гигантопирамидальных нейронов предцентральной извилины, ее верхних 2/3. Аксоны этих клеток направляются к внутренней капсуле, проходят через переднюю часть ее задней ножки (сразу позади волокон корково-ядерного пути), спускаются в основание ножки мозга, где занимают место латеральнее корково-ядерного пути.
Далее корково-спинномозговые волокна спускаются в переднюю часть (основание) моста, пронизывают идущие в поперечном направлении пучки волокон моста и выходят в продолговатый мозг, где на передней (нижней) его поверхности образуют выступающие вперед валики - пирамиды.
В нижней части продолговатого мозга часть волокон переходит на противоположную сторону и продолжается в боковой канатик спинного мозга, постепенно заканчиваясь в передних рогах спинного мозга синапсами на двигательных клетках его ядер.
Эта часть пирамидных путей, участвующая в образовании перекреста пирамид (моторный перекрест), получила название латерального корково-спинномозгового пути. Те волокна корково-спинномозгового пути, которые не участвуют в образовании перекреста пирамид и не переходят на противоположную сторону, продолжают свой путь вниз в составе переднего канатика спинного мозга. Эти волокна составляют передний корково-спинномозговой путь.
Затем эти волокна также переходят на противоположную сторону, но через белую спайку спинного мозга и заканчиваются на двигательных клетках переднего рога противоположной стороны спинного мозга. Располагающийся в переднем канатике передний корково-спинномозговой путь более молодой в эволюционном плане, чем латеральный. Его волокна спускаются преимущественно до уровня шейных и грудных сегментов спинного мозга.
Следует отметить, что все пирамидные пути являются перекрещенными, т.е. их волокна на пути к следующему нейрону рано или поздно переходят на противоположную сторону. Поэтому повреждение волокон пирамидных путей при одностороннем поражении спинного (или головного) мозга ведет к параличу мышц на противоположной стороне, получающих иннервацию из сегментов, лежащих ниже места повреждения.
Вторыми нейронами нисходящего произвольного двигательного пути (корково-спинномозгового) являются клетки передних рогов спинного мозга, длинные отростки которых выходят из спинного мозга в составе передних корешков и направляются в составе спинномозговых нервов для иннервации скелетных мышц.
Экстрапирамидные проводящие пути, объединенные в одну группу, в отличие от более новых пирамидных путей являются эволюционно более старыми, имеющими обширные связи в мозговом стволе и с корой большого мозга, взявшей на себя функции контроля и управления экстрапирамидной системой.

Кора большого мозга, получающая импульсы как по прямым (коркового направления) восходящим чувствительным путям, так и из подкорковых центров, управляет двигательными функциями организма через экстрапирамидные и пирамидные пути. Кора большого мозга оказывает влияние на двигательные функции спинного мозга через систему мозжечок - красные ядра, через ретикулярную формацию, имеющую связи с таламусом и полосатым телом, через вестибулярные ядра.
Таким образом, в число центров экстрапирамидной системы входят красные ядра, одной из функций которых является поддержание мышечного тонуса, необходимого для удерживания тела в состоянии равновесия без усилия воли. Красные ядра, которые относятся также к ретикулярной формации, получают импульсы из коры большого мозга, мозжечка (от мозжечковых проприоцептивных путей) и сами имеют связи с двигательными ядрами передних рогов спинного мозга.
Красноядерно-спинномозговой путь (trdctus rubrospinalis) входит в состав рефлекторной дуги, приносящим звеном которой являются спинно-мозжечковые проприоцептивные проводящие пути. Этот путь берет начало от красного ядра (пучок Монакова), переходит на противоположную сторону (перекрест Фореля) и спускается в боковом канатике спинного мозга, заканчиваясь на двигательных клетках спинного мозга. Волокна этого пути проходят в задней части (покрышка) моста и боковых отделах продолговатого мозга.
Важным звеном в координации двигательных функций тела человека является преддверно-спинномозговой путь (tractus vestibulospinalis).
Он связывает ядра вестибулярного аппарата с передними рогами спинного мозга и обеспечивает установочные реакции тела при нарушении равновесия. В образовании преддверно-спинномозгового пути принимают участие аксоны клеток латерального вестибулярного ядра (ядро Дейтерса), а также нижнего вестибулярного ядра (нисходящего корешка) преддверно-улиткового нерва.
Эти волокна спускаются в латеральной части переднего канатика спинного мозга (на границе с боковым) и заканчиваются на двигательных клетках передних рогов спинного мозга. Ядра, образующие преддверно-спинномозговой путь, находятся в непосредственной связи с мозжечком, а также с задним продольным пучком (fasciculus longitudinalis dorsalis, s. posterior), который в свою очередь связан с ядрами глазодвигательных нервов.
Наличие связей с ядрами глазодвигательных нервов обеспечивает сохранение положения глазных яблок (направление зрительной оси) при поворотах головы и шеи.
В образовании заднего продольного пучка и тех волокон, которые достигают передних рогов спинного мозга (ретикулярно-спинномозговой путь, tractus reticulospinalis), принимают участие клеточные скопления ретикулярной формации стволовой части мозга, главным образом промежуточное ядро (nucleus intersticialis, ядро Кахаля), ядро эпиталамической (задней) спайки, ядро Даркшевича, к которым приходят волокна из базальных ядер полушарий большого мозга.
Управление функциями мозжечка, участвующего в координации движений головы, туловища и конечностей и связанного в свою очередь с красными ядрами и вестибулярным аппаратом, осуществляется из коры большого мозга через мост по корково-мостомозжечковому пути (tractus corticopontocerebellaris). Этот проводящий путь состоит из двух нейронов. Тела клеток первого нейрона лежат в коре лобной, височной, теменной и затылочной долей. Их отростки - корковом остовые волокна (fibrae corticopontinae) направляются к внутренней капсуле и проходят через нее. Волокна из лобной доли, которые можно назвать лобно-мостовыми волокнами (fibrae frontopontinae), проходят через переднюю ножку внутренней капсулы. Нервные волокна из височной, теменной и затылочной долей идут через заднюю ножку внутренней капсулы.
Далее волокна корково-мостового пути идут через основание ножки мозга. От лобной доли волокна проходят через самую медиальную часть основания ножки мозга, кнутри от корково-ядерных волокон.
От теменной и других долей полушарий большого мозга идут через самую латеральную часть, кнаружи от корково-спинномозговых путей. В передней части (в основании) моста волокна корково-мостового пути заканчиваются синапсами на клетках ядра моста этой же стороны мозга. Клетки ядер моста с их отростками составляют второй нейрон корково-мозжечкового пути.
Аксоны клеток ядер моста складываются в пучки - поперечные волокна моста (fibrae pontis transversae), которые переходят на противоположную сторону, пересекают при этом в поперечном направлении нисходящие пучки волокон пирамидных путей и через среднюю мозжечковую ножку направляются в полушарие мозжечка противоположной стороны.
Таким образом, проводящие пути головного и спинного мозга устанавливают связи между афферентными и эфферентными (эффекторными) центрами, участвуют в образовании сложных рефлекторных дуг в теле человека.
Одни проводящие пути (системы волокон) начинаются или заканчиваются в эволюционно более старых, лежащих в мозговом стволе ядрах, обеспечивающих функции, обладающие определенным автоматизмом. Эти функции (например, тонус мышц, автоматические рефлекторные движения) осуществляются без участия сознания, хотя и под контролем коры большого мозга.
Другие проводящие пути передают импульсы в кору большого мозга, в высшие отделы ЦНС, или из коры к подкорковым центрам (к базальным ядрам, ядрам мозгового ствола и спинного мозга).