Тема лекции
Все живые клетки отделены от окружающей среды поверхностью, называемой клеточной мембраной. Кроме того, для эукариотов характерно образование внутри клеток компартментов, то есть деление клетки на определенные части, которые отделены внутриклеточными мембранами от цитоплазмы и выполняющая свою определенную функцию. По другому компартменты называют субклеточными органеллами, выделяют ядро, митохондрии, комплекс Гольджи, эндоплазматическая сеть, пероксисомы, лизосомы и т.д. Мембраны представляют собой не только статически организованные поверхности раздела, но и включают активные биохимические системы, отвечающие за такие процессы, как избирательный транспорт веществ внутрь и наружу клетки, связывание гормонов и других регуляторных молекул, протекание ферментативных реакций, передача импульсов нервной системы и т.д. (рис. 1).

Рис. 1 Функции биомембран
Мембраны состоят из липидных и белковых молекул, относительное количество которых варьирует (от 1/5 — белок + 4/5 — липиды до 3/4 — белок + 1/4 – липиды) у разных мембран. Углеводы содержатся в форме гликопротеинов, гликолипидов и составляют 0,5–10% вещества мембраны.
Основная часть липидов в мембранах представлена фосфолипидами, гликолипидами и холестеролом. Строение этих липидов представлено на рис. 2. Липиды мембран имеют в структуре две части: неполярный гидрофобный «хвост» и полярную гидрофильную «голову». Такую двойственную природу соединений называют амфифильной. Липиды мембран образуют двухслойную структуру. Каждый слой состоит из сложных молекул липидов, расположенных таким образом, что неполярные гидрофобные «хвосты» находятся в тесном контакте друг с другом. Так же контактируют гидрофильные части молекул. Все взаимодействия имеют нековалентный характер. Два монослоя ориентируются «хвост к хвосту», так, что образующаяся структура двойного слоя имеет внутреннюю неполярную часть и две полярные поверхности. Белки мембран включены в липидный бислой тремя способами:
1. связаны с гидрофильной поверхностью липидного бислоя — поверхностные мембранные белки;
2. погружены в гидрофобную область бислоя;
3. пронизывают мембрану — интегральные мембранные белки.

Рис. 2 Структура некоторых важнейших классов липидов, формирующих мембраны.

Рис. 3 Структура некоторых важнейших классов липидов, формирующих мембраны.
Поверхностные белки своими гидрофильными радикалами аминокислот взаимодействуют нековалентными связями с гидрофильными группами липидного бислоя. Интегральные белки различаются по степени погруженности в гидрофобную часть бислоя липидов. Они могут располагаться внутри мембраны и частично погружаться в мембрану либо прошивать мембрану насквозь. Внутримембранная часть интегральных белков содержит большое количество аминокислот с гидрофобными радикалами, которые обеспечивают гидрофобное взаимодействие с липидами мембран. Гидрофобные взаимодействия поддерживают определенную ориентацию белков в мембране. Гидрофильная выступающая часть белка не может переместиться в гидрофобный слой. Часть мембранных белков ковалентно связаны с моносахаридными остатками или олигосахаридными цепями и представляют собой гликопротеины. Примеры расположения белков и липидов в мембране представлены на рис. 3.
Хотя каждый монослой образован из липидов, ориентированных одинаковым образом, тем не менее липидный состав монослоев различен. Например, в плазматической мембране эритроцитов фосфатидилхолины преобладают в наружном слое, а фосфатидилсерины — во внутреннем слое мембраны. Углеводные части белков и липидов располагаются на наружной части мембраны. Кроме того, поверхности мембраны отличаются по составу белков. Степень такой асимметрии мембран различна у разных типов мембран и может меняться в процессе жизнедеятельности клетки и ее старения. Жесткость и текучесть мембран также зависят от ее состава. Повышенная жесткость обуславливается увеличением соотношения насыщенных к ненасыщенным жирным кислотам в составе фосфолипидов, а также холестерола. Физические свойства мембран зависят от расположения белков в липидном слое. Липиды мембран способны к диффузии в пределах слоя параллельно поверхности мембраны (латеральная диффузия). Поверхностные белки тоже способны к латеральной диффузии. Поперечная диффузия в мембранах сильно ограничена. Таким образом, модель, которая отображена на рис. 3 объясняет ряд экспериментальных фактов и называется жидко-кристаллической, то есть подвижной системой, имеющей определенную упорядоченность слегка сходную с кристаллической структурой.
Мембранный транспорт
Транспорт веществ внутрь и наружу клетки, а также между цитоплазмой и различными субклеточными органеллами (митохондриями, ядром и т.д.) обеспечивается мембранами. Если бы мембраны были глухим барьером, то внутриклеточное пространство оказалось бы недоступным для питательных веществ, а продукты жизнедеятельности не могли бы быть удалены из клетки. В то же время при полной проницаемости было бы невозможно накопление определенных веществ в клетке. Транспортные свойства мембраны характеризуются полупроницаемостью: некоторые соединения могут проникать через нее, а другие — нет (рис. 4). Одна из главных функций мембран — регуляция переноса веществ. Существуют два способа переноса веществ через мембрану: пассивный (А) и активный (Б) транспорт (рис. 5).
Пассивный транспорт (или облегченная диффузия). Если вещество движется через мембрану из области с высокой концентрацией в сторону низкой концентрации (т.е. по градиенту концентрации этого вещества) без затраты клеткой энергии, то такой транспорт называется пассивным, или диффузией. Различают два типа диффузии: простую (неспецифическую) и облегченную. Простая диффузия характерна для небольших нейтральных молекул (H2O, CO2, O2), а также гидрофобных низкомолекулярных органических веществ. Эти молекулы могут проходить без какого-либо взаимодействия с мембранными белками через поры или каналы мембраны до тех пор, пока будет сохраняться градиент концентрации (рис. 4).
Облегченная диффузия характерна для гидрофильных молекул, которые переносятся через мембрану также по градиенту концентрации, но с помощью специальных мембранных белков-переносчиков (рис. 4). Для облегченной диффузии в отличие от простой диффузии характерна высокая избирательность, так как белок-переносчик имеет центр связывания, комплементарный транспортируемому веществу, и перенос сопровождается конформационными изменениями белка.

Рис. 4 Селективная проницаемость мембран по отношению к веществам.
Один из возможных механизмов облегченной диффузии может быть следующим: транспортный белок (транслоказа) связывает вещество, затем сближается с противоположной стороной мембраны, освобождает это вещество, принимает исходную конформацию и вновь готов выполнять транспортную функцию. Малоизвестно о том, как осуществляется передвижение самого белка. Другой возможный механизм переноса предполагает участие нескольких белков-переносчиков. В этом случае первоначально связанное соединение само переходит от одного белка к другому, последовательно связываясь то с одним, то с другим белком, пока не окажется на противоположной стороне мембраны.

Рис. 5 Виды транспортных процессов – энергозависимые и энергонезависимые.
Активный транспорт имеет место в том случае, когда перенос осуществляется против градиента концентрации. Такой перенос требует затраты энергии клеткой. Активный транспорт служит для накопления веществ внутри клетки. Источником энергии часто является АТФ. Для активного транспорта кроме источника энергии необходимо участие мембранных белков. Одна из активных транспортных систем в клетке животных отвечает за перенос ионов Na+ и K+ через клеточную мембрану. Эта система называется Na+, K+-насос. Она отвечает за поддержание состава внутриклеточной среды, в которой концентрация K+ выше, чем Na+ (рис. 5). Градиент концентрации калия и натрия поддерживается путем переноса К+ внутрь клетки, а Na+ наружу. Оба транспорта происходят против градиента концентрации. Такое распределение ионов определяет содержание воды в клетках, возбудимость нервных клеток и клеток мышц и другие свойства нормальных клеток. Na+, K+ –насос представляет собой белок — транспортную АТФ-азу. Молекула этого фермента является олигомером и пронизывает мембрану. За полный цикл работы насоса из клетки в межклеточное вещество переносятся три иона Na+, а в обратном направлении — два иона К+. При этом используется энергия молекулы АТФ. Транспортная АТФаза в этом случае имеет центры связывания для обоих веществ (рис. 6). Существуют транспортные системы для переноса ионов кальция (Са2+- АТФ-азы), протонные насосы (Н+-АТФазы) и др. Нередко перенос веществ осуществляется по механизму вторичного активного транспорта за счет симпорта или антипорта двух веществ. Симпорт — это активный перенос вещества через мембрану, осуществляемый за счет энергии градиента концентрации другого вещества, который движется по градиенту концентрации. Антипорт — это перемещение вещества против градиента своей концентрации. При этом другое вещество движется в противоположном направлении по градиенту концентрации. Симпорт и антипорт могут происходить при всасывании аминокислот из кишечника и реабсорбции глюкозы из первичной мочи. При этом используется энергия градиента концентрации ионов Na+, создаваемого Na+,K+- АТФ-азой.
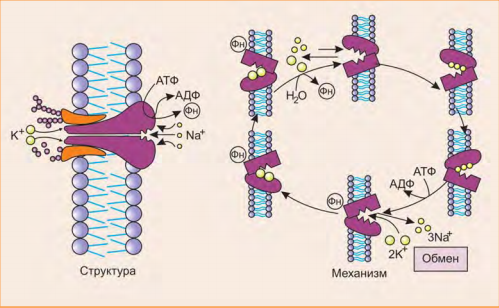
Рис. 6 Механизмы реализации активного транспорта.