Глава 15
ЭКОЛОГИЧЕСКИЕ АСПЕКТЫ
ПАРАЗИТОЛОГИИ
Паразитология — наука экологического профиля, поскольку паразитизм является одной из форм межвидовых отношений. Вместе с тем экологическим аспектам паразитологии (расселение паразитов, поиски хозяина, взаимоотношения паразита и хозяина) уделяется меньше внимания, чем морфологии, систематике паразитов, изучению их циклов развития и т. д.
Среди советских паразитологов вопросами экологии паразитО! занимались Е. Н. Павловский, К. И. Скрябин, В. Н. Беклемишев и основатель советской экологической школы паразитологов В. А. Догель, труды которых внесли существенный вклад в развитие этого направления.
ВЗАИМООТНОШЕНИЯ В СИСТЕМЕ
ПАРАЗИТ — ХОЗЯИН НА УРОВНЕ ПОПУЛЯЦИЙ
Для современной экологической паразитологии характерен переход 1Н изучения отношений в системе паразит — хозяин на уровне шдельных особей к изучению этого вопроса на уровне II О II у л я ц и й.
Развивающаяся популяционная экология паразитов уже накопила ишересный фактический материал, позволяющий дать ответ на ряд мопросов, остававшихся нерешенными.
15.2. ЗНАЧЕНИЕ ПАРАЗИТИЗМА КАК ФАКТОРА
УСТОЙЧИВОСТИ СИСТЕМЫПАРАЗИТ - ХОЗЯИН
Подход к изучению проблемы паразитизма на популяционном уровне позволил выявить ряд новых закономерностей, среди которых важно отметить неодинаковую биологическую значимость паразитизма пик отдельных особей хозяина и для популяции хозяина.
На уровне отдельных особей паразит, как правило, приносит вред организму хозяина, ослабляет его жизнедеятельность иногда даже убивая его. В популяции хозяина гибель части особей под воздействием паразитов оказывается не вредной, а «полезной» для этой группы организмов. Например, при возрастании скорости размножения количество особей в популяции хозяина может превысить пищевые |ч-сурсы, имеющиеся в ее ареале, и вызвать самоуничтожение популяции. Благодаря гибели части животных под воздействием паразитов вновь восстанавливается оптимальное соотношение между числом особей популяции и конкретными возможностями среды обитания. В результате популяция хозяина сохраняется, т. е. ее устойчивость повышается. Одновременно выживает и популяция паразита. Следовательно, паразитизм, регулируя численность популяции хозяина, повышает устойчивость системы паразит—хозяин.
Этот регулирующий механизм вступает в действие при определенных условиях — бескормица, авитаминоз, повышение плотности популяции и др. Главную роль при этом играют наиболее патогенные паразиты, вызывающие массовые острые забопевания животных или шизоотии.
Популяции хозяина являются составным компонентом различных биогеоценозов. При резком повышении их плотности происходит разрушение нижележащих трофических уровней биоценоза, что нарушает равновесие между элементами экосистемы. Так, при резком увеличении численности насекомых, гусеницы которых питаются листьями, последние могут уничтожить листву деревьев полностью, вызванная дефолиация означает разрушение первичного трофического уровня, что имеет тяжелые последствия для всех элементов экосистемы. Таким образом, согласно новым данным, паразитизм следует рассматривать как экологический фактор, который регулирует численность популяции хозяина и повышает устойчивость не только простых систем паразит—хозяин на уровне популяции, но и экосистем боле< сложного уровня — биоценозов.
С этой точки зрения представление о паразитарном заболевании ка)| отрицательном явлении по отношению к природным биоценозам должно быть пересмотрено как не соответствующее сущности явления (В. Н. Беклемишев). Однако следует сразу отметить, что подобное! значение паразитизма ни в коей мере не распространяется на популяции современного человечества.
Новые представления о «полезной» роли паразитизма как возможного универсального фактора, регулирующего устойчивость экосистем, позволили объяснить широкое распространение парази>; тизма и относительную устойчивость систем паразит—хозяин.
Получил объяснение и другой вопрос, а именно почему у хозяев паразитов, принадлежащих к высокоорганизованным группам животного мира и обладающих хорошо сформированными защитными реакциями, в процессе эволюции не выработалась защита от паразитизма. Стало очевидно, что подобные приспособления не могли закрепляться в процессе естественного отбора. В освободившихся от, паразитов популяциях хозяев численность особей стала бы возрастать,' что привело бы к быстрому разрушению элементов среды, необходи-1 мой для популяции, в частности к истощению пищевых ресурсов и \ жизненного пространства. По данным некоторых авторов у млекопи-' тающих, наоборот, обнаружены приспособления, облегчающие парази-; тизм. }
Современный регулирующий численность популяций хозяев меха-! низм паразитизма является результатом длительной эволюции экоси-] стем. В соответствии с разнообразием паразитов, их циклов и парази- ] тических связей, паразитизм представляет собой гибкий и быстро{ реагирующий фактор экосистем. 1
Как уже указывалось выше, по отношению к популяциям человека в настоящее время паразитизм не играет положительной роли, хотя, безусловно, биологическая эволюция человека также контролировалась паразитами, поддерживавшими численность популяций людей в соответствии с экологическими возможностями среды. Считают, что на ранних этапах цивилизации рост популяции человечества еще контролировался паразитарным регулирующим механизмом и возникавшие эпидемии вызывали гибель больших масс людей. Однако с развитием хозяйственной деятельности человека и увеличением производства продуктов питания численность популяций человека стала расти, что привело к нарушениям устойчивости системы. На более поздних этапах цивилизации обществу пришлось организовывать защиту от действия паразитарного механизма и разработать для этого систему лечебных и профилактических мероприятий.
Однако в развивающихся странах, особенно в тропических, до настоящего времени паразитарные заболевания служат причиной высокой смертности среди населения. По данным ВОЗ, около 400 млн. человек поражены гельминтами, в связи с чем ежегодно погибают свыше 1 млн. детей.
Человек научился использовать паразитический механизм в своих целях. Искусственно разводя паразитических насекомых, человек затем использует их для подавления резких вспышек численности насекомых- нредителей.
15.3.РАСПРЕДЕЛЕНИЕ ПАРАЗИТОВ В ПОПУЛЯЦИИ ХОЗЯИНА
В пределах одной популяции степень зараженности разных особей неодинакова. Как показали исследования, на уровне популяций распределение паразитов между особями хозяина соответствует не случайному или нормальному, а так называемому перерассеянному шну распределения, т. е. паразиты сосредоточены лишь в немногих особях хозяина, но в большом количестве.
Наиболее частая причина неоднородности заражения особей хозяина мключается в различной вероятности инвазии для различных особей хозяина и различных особей паразита. Для того чтобы распределение паразита в популяции хозяина было случайным, вероятность инвазии должна быть одинаковой для всех особей хозяина и всех особей паразита, т. е. среда должна быть однородной. Поскольку в природе такие условия отсутствуют, возникают отклонения в концентрации паразитов II отдельных особях.
Второй причиной перерассеянного распределения паразита является возможность бесполого или партеногенетического размножения пара- шта в организме хозяина (малярийный плазмодий в человеке, фематоды в промежуточных хозяевах и др.).
И, наконец, суперинвазия становится возможной вследствие ослабления защитной реакции хозяина, что повышает вероятность его последующих повторных заражений.
Перерассеянное распределение паразита в популяции необходимо учитывать при разработке мер борьбы и профилактики паразитарных (аболеваний.
15.4.РАССЕЛЕНИЕ И ПОИСК ХОЗЯИНА
Расселение животных необходимо для избежания перенаселенности, повышающей смертность, и поэтому служит важным условием сохранения вида. Особенно важно расселение для паразитов, поскольку су пер инвазия приводит к смерти хозяина и гибели паразита. Поэтому паразиты обладают способностью покидать на определенной стадии развития хозяина и находить новых хозяев.
Переход от одного хозяина к другому, обеспечивая расселение паразита, одновременно влечет за собой высокую его смертность, поскольку вероятность нахождения нового хозяина, как правило, крайне мала. Гибель большого числа особей на разных стадиях цикла развития паразита компенсируется высокой плодовитостью паразитических форм.
Расселение паразитов происходит различными способами и на различных стадиях жизненного цикла..
Наиболее часто переход от одного хозяина к другому осуществляется с помощью свободноживущей (находится вне организма хозяина) активной подвижной (личинка) и неактивной покоящейся (яйцо, цисте спора) стадий. Последняя имеет защитные приспособления (оболочкам обеспечивающие переживание неблагоприятных условий, и обычнв неподвижна. Благодаря этому она не может обеспечить расселение пространстве, но позволяет продлить срок существования паразита! т. е. обеспечивает его расселение во времени, что повышает шансы на встречу с новым хозяином.
Активная подвижная стадия (церкарий, рабдитная, филяриевидная личинка) способствует расселению паразита в пространстве, обычно ни питается и живет недолго. Кроме функции расселения, она выполняет it функции поиска хозяина.
Включение в цикл развития промежуточного хозяина дает возможность паразиту избежать неблагоприятных воздействий среды и продлить срок своего существования. ^
Расселению паразита способствует также перемещение хозяев. Особенно эффективно перемещение окончательного хозяина, содержа»| щего половозрелые формы и рассеивающего яйца по территории! обитания.
Если цикл развития паразита включает переносчика, то функцию поиска нового хозяина выполняет переносчик. При наличии в цикле1 промежуточного хозяина паразит, как правило, не участвует в поиске окончательного хозяина, а попадает к нему благодаря существующим пищевым связям.,
Паразиты могут изменять поведение промежуточных хозяев таким образом, что шансы попасть к окончательному хозяину будут повышаться. Например, муравьи, зараженные метацеркариями трема» тоды, выползают на кончик травинки и здесь впадают в состояние свое«образного оцепенения, что обеспечивает заглатывание их травоядными животными. Рыбы, зараженные некоторыми видами цестод, значительно медленнее плавают, отчего становятся добычей хищников • первую очередь.
У многих паразитов вероятность встречи с хозяином облегчается за счет синхронизации их цикла развития с циклом развития хозяина или переносчика. Так, у трематоды, паразитирующей в мочевом пузыре лягушки, яйца созревают и выделяются в тот период, когда лягушки переходят в воду для размножения (продолжительность периода почти неделя). Из отложенных яиц паразита инвазионные личинки выходят а тот момент, когда у головастика появляются. жабры, на которых личинки паразитируют. Таким образом, и инвазионные стадии, и хозяева находятся одновременно в одном месте, синхронность циклов развития паразита и хозяина вызывается одновременным воздействием гипофизарного гормона лягушки и на организм паразита.
Яйца Schistosoma haematobium выделяются из организма человека с мочой, причем наибольшее их количество поступает в полость мочевого пузыря в полдень, когда вероятность купания особенно велика (промежуточный хозяин — водный моллюск).
Примером синхронизации циклов паразита и переносчика может служить филярия Wuchereria bankrofti, личинки которой поступают в периферические сосуды человека в ночные часы, т. е. в то время, когда переносчик комар рода Mansonia нападает на человека для сосания крови. В дневные часы личинки концентрируются в сосудах внутренних органов.
ПРИНЦИПЫСИСТЕМАТИКИ
Объектом медицинской паразитологии, как уже отмечалось, служат паразиты и переносчики, принадлежащие к различным группам животного мира. Необходимой предпосылкой их изучения является знание основ систематики или классификации животного мира.
Систематика — наука, подразделяющая животный и растительный мир на соподчиненные группы в зависимости от их сходства или различия, и строящая систему животного мира. Возникновение систематики было вызвано необходимостью ориентироваться в многообразии животных форм. Первоначально ее задача была чисто практической и состояла в выделении полезных и вредных для человека организмов (съедобные, несъедобные, ядовитые, неядовитые). С момента появления теории Ч. Дарвина и до настоящего времени основной «дачей систематики является отражение исторического родства существующих форм.
Принципы систематики были разработаны шведским естествоиспытателем К. Линнеем (1707—1778) и изложены в работе «Система природы» (1735). К. Линней выделил четыре систематические категории: вид, род, отряд и класс. Вид, по К. Линнею, наименьшая система- I ическая единица, объединяющая организмы, наиболее сходные между собой. Сходные виды объединяются в род, роды — в отряд, отряды — в класс, который представлял высшую систематическую единицу. Весь животный мир, по К. Линнею, включал всего 6 классов: млекопитающие, птицы, гады, рыбы, насекомые и черви.
Система К. Линнея была искусственной, поскольку не отражала реального исторического родства между организмами, так как в основу деления на группы было положено формальное сходство отдельных признаков.
Многие классификации, создававшиеся в последующие годы, также были искусственными или в значительной мере формальными. Современная система животного мира является результатом длительного развития биологических наук. Она естественна и отражает реальные филогенетические связи между группами организмов, поскольку в основу классификации положена совокупность признаков организмов.
Система соподчиненных групп современной классификации включает вид (species), род (genus), семейство (familia), отряд (ordo), класс (classis) и тип (typus). Для некоторых групп применяют промежуточные Фуппы, подтипы, подотряды, подсемейства. Количество видов в животном мире составляет более 2 млн., типов — свыше 20.
В систематике принята двойная, или бинарная, номенклатура, впервые введенная К. Линнеем. Сущность бинарной номенклатуры состоит в том, что каждый вид имеет двойное наименование. Первое слово означает название рода, к которому относится вид, второе -в собственно видовое название. Например, клещ таежный — Ixodei persulcatus, клещ собачий — Ixodes ricinus (слово Ixodes означает! название рода, а persulcatus и ricinus — название видов). Бинарная номенклатура представляет общепринятую международную систему, в] которой используется только латинская терминология. Благодаря этому возможно взаимопонимание специалистов любых стран. '
Врачу необходимо знать систематическое положение важнейших паразитов человека и уметь определять их видовую принадлежность, поскольку каждый вид обладает своими особенностями строения И экологии. Без этого невозможно поставить точный клинический] диагноз и обосновать соответствующие меры профилактики. j
Глава 16
ПРОСТЕЙШИЕ
(МЕДИЦИНСКАЯ ПРОТОЗООЛОГИЯ)
К типу простейших (Protozoa) относится ряд патогенных для человека форм, поражающих отдельные ткани и органы и вызывающих заболевания различной тяжести, в том числе и с летальным (смертельным) исходом.
Наука о паразитических простейших — возбудителях болезней человека — носит название протозоология и является важным разделом в подготовке врачей разных специальностей.
Морфофизиологическая характеристика. По строению соответствуют отдельной клетке многоклеточных (отсюда название «одноклеточные»), по функциям — целому самостоятельному организму. Принципиальным отличием простейших от клетки многоклеточных служит специализация последних, т. с. каждый вид клеток выполняет в организме какую-нибудь одну функцию и потому зависит от деятельности других клеток и не может существовать изолированно. В противоположность этому единственная клетка, которая представляет собой организм простейших, передвигается, захватывает пищу, размножается, защищается от врагов, т. е. обладает всеми свойствами целого организма и физиологически ему соответствует. Поэтому в настоящее время простейших называют организмами на клеточном уровне. В системе животного мире простейшие представлены всего одним типом.
Тело простейших имеет микроскопические размеры и состоит из тех же компонентов, что и клетка многоклеточных — наружной мембраны, цитоплазмы, ядра и органоидов.
Наружная мембрана имеет типичное трехслойное строение, но за счет дополнительных образований у некоторых видов может достигать большой толщины и плотности. Цитоплазма подразделяется на два слоя: наружный и внутренний. Наружный слой (эктоплазма) более плотный, однородный и прозрачный, внутренний (эндоплазма) — зернистый, имеет более жидкую консистенцию. В эндоплазме нахо«дятся органоиды общего назначения — митохондрии, эндоплазматическая сеть, сетчатый аппарат и др. Кроме того, в соответствии с функциями, присущими целому организму, простейшие имеют органоиды специального назначения, осуществляющие функции передвижения, ми гания, выделения, защиты и др.
Органоидами движения простейших служат: 1) псевдоподии или ножноножки, представляющие собой временные выросты цитоплазмы;
жгутики— постоянные органоиды, имеющие вид длинной тонкой мши, начинающейся обычно на переднем конце; 3) реснички — постоянные органоиды, представляющие собой многочисленные короткие нити.
Строение органоидов питания не одинаково и зависит от способа питания разных простейших. Большая часть простейших питается частицами твердой пищи. У таких организмов для переваривания пищи существует пищеварительная вакуоль — капля жидкости, содержащая пищеварительные ферменты, которая образуется при поступлении нищи в эндоплазму. Пищеварительная вакуоль окружает пищевую частицу и перемещается по телу простейшего. Пища переваривается и всасывается в цитоплазму. Остатки непереваренной пищи вместе с пищеварительной вакуолью выбрасываются наружу.
Многие простейшие, ведущие паразитический образ жизни, усваивают пищу всей поверхностью тела, используя в основном механизм пиноцитоза. Наконец, небольшая группа простейших питается подобно растениям и имеет хлоропласты.
Органоиды выделения представлены сократительной или пульсирующей вакуолью, имеющей вид небольшого пузырька, наполненного жидкостью, который периодически увеличивается, достигая определенного объема, а затем сокращается, выбрасывая жидкость наружу. Накопление и сокращение ритмически чередуются. Основная функция сократительной вакуоли — поддержание осмотического давления на постоянном уровне. Концентрация минеральных и органических веществ, определяющая осмотическое давление, в теле простейших выше, чем в окружающей среде, поэтому вода по закону осмоса[1] постоянно поступает в цитоплазму. Если воду не удалять, то произойдет набухание и гибель простейшего.
У паразитических и морских форм, обитающих в среде с более высоким осмотическим давлением, сократительная вакуоль может отсутствовать.
Одновременно с излишками воды через сократительную вакуоль удаляются жидкие продукты диссимиляции и осуществляется снабжение клетки кислородом, поступающим с водой.
Органоиды защиты см. 16. 4.
Большинство простейших имеет одно ядро, но существуют и многоядерные формы. Ядро имеет строение, характерное для »укариот. Морфология и формы деления ядер разнообразны, но, как доказано в настоящее время, деление ядра в любом случае представляв' собой митоз.
Размножение. Простейшие могут размножаться бесполым I половым путем. Бесполое размножение встречается как в виде деленй на две части, так и в виде множественного деления. Половой процесс ] большинства простейших представлен копуляцией, у инфузорий -1 конъюгацией.
Жизненный цикл. В отличие от соматических клето* многоклеточных, жизненный цикл простейших складывается из, последовательных стадий развития, которые повторяются с опреде- ленной закономерностью. Чаще всего исходной стадией является; зигота, затем следует бесполое размножение путем деления, затем образование половых особей и вновь образование зиготы.
Инцистйрование. При неблагоприятных условиях внешней среды (повышение или понижение температуры, высыхание и т. д.) вегетативные формы простейших превращаются в цисты. Они перестают питаться, двигаться и покрываются толстой оболочкой; процессы обмена веществ при этом резко замедляются. При, восстановлении благоприятных условий из цисты вновь выходит активная вегетативная форма. Инцистирование — защитная реакция, возникшая в процессе эволюции, обеспечивающая выживание в неблагоприятных условиях. Цисты могут сохраняться в течение длительного времени, месяцы и даже годы.
Распространение. Простейшие обитают в пресной и морской воде, жидких средах организмов.
Классификация. В настоящее время классификация тип простейших пересматривается. Появились новые классификации. Некоторые авторы (В. А. Догель) выделяют простейших как царство! которое делится на 5 самостоятельных типов: саркомастигофора, споровики, книдоспоридии, микроспоридии и инфузории. Общепринятой является классификация, согласно которой все простейшие делятся на 4 класса: 1. Класс саркодовые; 2. Класс жгутиковые; 3. Класс споровики; 4. Класс инфузории;
Паразиты человека встречаются в каждом классе!
16.1. КЛАСС САРКОДОВЫЕ (5А1<СООША)
Морфофизиологическая характеристика. Наиболее примитивные простейшие, что проявляются прежде всего §' слабой степени дифференцировки. Цитоплазма ограничена только наружной мембраной, форма тела непостоянная. Органоидами движО" кия и захвата пищи служат псевдоподии. Специальное ротово! отверстие отсутствует. Поступление пищи и выделение непереваренных остатков может происходить в любом участке тела. Сократительная вакуоль одна. Выделение продуктов диссимиляции и излишков воды также может происходить в любом месте. Обычно имеется только одно ядро, хотя встречаются и многоядерные формы.
Размножение. Размножение в основном бесполым путем —«делением на две части
Инцистирование. В неблагоприятных условиях образуют цисту.
Медицинское значение имеет только один отряд подкласса Корненожек, а именно отряд Амеб (Amoebina).
16.1.1. Отряд Амебы (Amoebina)
В организме человека обитает несколько видов амеб. Наиболее важное значение имеет дизентерийная амеба.
Дизентерийная амеба (Entamoeba histolytica). Возбудитель тяжелого 1аболевания — амебной дизентерии или амебиаза.
Локализация. Толстый кишечник, v
Географическое распространение. Повсеместно, но в районах с жарким климатом встречается чаще.


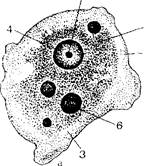
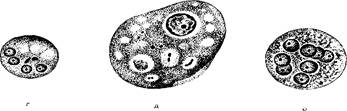
Рис. 172. Дизентерийная и кишечная амебы.
Чнаентерийная амеба: а крупная вегетативная форма; б, в - мелкая вегетативная форма: 1 - эктоплазма; 2 эндоплазма; 3 -- псевдоподии; 4 — ядро; 5 — кариоеома; 6 — фагоцитированные эритроциты п пищеварительных вакуолях; г — циста; кишечная амеба: А ~ - вегетативная форма; е -- циста.
Морфофизиологическая характеристика и
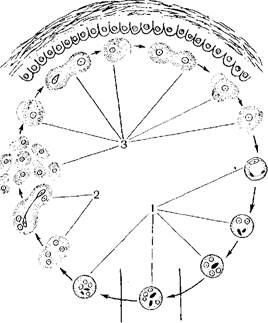
Рис. 173. Жизненный цикл дизентерийной амебы.
а - - взрослый здоровый носители] б ■ (юлвиой амебиазом; 1 цисти; 2 эксцис!ированная вегетативна» форма;.1 мелкая вегетативна» форма, крупная вечетативкая форм» 1 е заглоченными чритропита'.^ 5 в елнзиетои кишечника; 6 7 выделяющаяся е фекалиями наружу.
жизненный цикл. Паразитирует только у человека. В жизненном цикле встречаются следующие формы: циста, мелкая вегетативная форма (forma minuta), крупная вегетативная форма (forma magna) и тканевая.
Инвазионной стадией является циста, содержащая 4 ядра (отличительный видовой признак). В кишечнике человека оболочка цисты растворяется и из нее выходит четырехядерная амеба, которая быстро делится на 4 одноядерные м"елкие (7—15 мкм в диаметре) вегетативные формы (f. minuta).
Мелкая вегетативная форма обитает в просвете толстого кишечника, питается в основном бактериями, размножается и нв вызывает заболевания. При попадании в нижние отделы толстого кишечника она превращается в цисту, содержащую сначала одно ядро, которое в процессе созревания делится с образованием четырехядерноЙ цисты.
У некоторых людей при соответствующих условиях (охлажденив(перегревание, авитаминозы, нарушения диеты, гельминтозы) formt minuta проникает в стенки кишечника, где интенсивно размножается И вызывает поражение слизистой с образованием язв. При этом разрушаются стенки кровеносных сосудов и возникают кровотечения > полость кишечника.
Пр и по явлении амебных поражений кишечника мелкие вегетативны«
Рис 173 продолжение

формы, находящиеся в просвете кишечника, начинают превращаться в крупную вегетативную форму. Последняя характеризуется крупными размерами (30—40 мкм) и строением ядра: хроматин ядра образует радиальные структуры, строго в центре располагается крупная глыбка хроматина — кариосома, f- magna начинает питаться эритроцитами, т. е. становится эри трофагом (рис. 172). Характерны тупые широкие псевдоподии и передвижение" толчками.
Амебы, размножающиеся в тканях стенки кишечника, — тканевая форма, — попадая в просвет, по строению и размерам становятся сходными с крупной вегетативной формой, но не способны заглатывать эритроциты.
Внедрение амеб в слизистую кишечника и ее расплавление связывают с выделением паразитами веществ, растворяющих тканевые белки (протеолитические ферменты, гиалуронидаза и др.).
При лечении или нарастании защитной реакции организма крупная 1югетатт1вная-фосма_вновь превращается в мелкую, которая начинает ннцистироваться (рисТТГЗ). В поСлёдугощем-ияи-наступает выздоровление, или заболевание переходит в хроническую форму.
У некоторых зараженных людей мелкая вегетативная форма никогда не превращается в крупную. Таких людей называют цистоносителями. Они представляют собой большую опасность, так как служат источником заражения окружающих. За сутки один цистоноситель выделяет до 600 млн. цист. Щисто носител и подле жа^ выявлению и обязательному лечению. |
Единственный источник заболевания амебиазом — человек. Выделяющиеся с фекалием цисты загрязняют почву и воду. Поскольку фекалии нередко используют как удобрение, цисты попадают в огород и сад, где загрязняют овощи и фрукты. Цисты устойчивы к воздействию внешней среды. В кишечник попадают с немытыми овощами и фруктами, через некипяченую воду, грязные руки. Механичеа^^ ками_сду2к ат мухи, тараканы, за грязняю щие пищ у.
Патогенное д ей с тТвТГе^ Развивается тяжелое заболева-1 ние, основными симптомами которого служат: KpQB0T04j^Uie-язвы ^ кишечнш<е1_частый и жидкий стул (до 10—20 раз в сутки) с примесью кров^1Гслит иногдаТГОТфовёносньш сосудам дизентерийная амеба может заноситься в печень ^и другие^рганы, вызыв ая там образование абсцессов (очаговыё~ттагноения). При отсутствии лечения смертность достигад2т40%._
Лабораторная диагностика. Микроскопирование мазков^ фекалий. Востром, периоде в мазке находятся крупные, вегетативные^Бо ^мьг, содерж ащие эритроцит ы^; цисты обычно отсутствуют, так как f. magna неспособна инцистироваться. При хрониче-; ской форме или цистоносительстве в фекалиях обнаруживаются четы- рех ядерные цисты. *""
П р о'ф илакт и к а: личная — обмывание овощей и фруктов кипяченой водой, употребление для питья только кипяченой воды, мытье рук перед едой, после посещения туалета и т. д.; общественная — борьба с загрязнением почвы и воды фекалиями, уничтожение мух, санитарно-просветительная работа, обследование на цистоноси- тельство лиц, работающих на предприятиях общественного питания, ] лечение больных. j
Наряду с дизентерийной амебой в пищеварительном тракте человекаг обитают саркодовые непатогенные или патогенность которых недостаточно доказана. Знание морфофизиологических особенностей этих амеб, необходимо, поскольку некоторые из них обладают сходством с дизентерийной амебой и могут быть приняты за патогенную форму.; В связи с этим врачу в ряде случаев приходится проводить дифференциальный диагноз между патогенными и непатогенными видами амеб. К числу непатогенных амеб относятся кишечная и ротовая амебы., Кишечная амеба (Entamoeba coli).
Локализация. Верхний отдел толстой кишки, обитает только в просвете кишечника.
Г еографическое распространение. Обнаруживается примерно у 40—50% населения различных областей земного шара.
Морфофизиологическая характеристика. Be-• гетативная форма имеет размеры 20—40 мкм, но иногда встречаются и более крупные формы. Резкая граница между экто- и эндоплазмой отсутствует. Обладает характерным способом передвижения — одновременно выпускает псевдоподии с разных сторон и как бы «топчется на месте». Ядро содержит крупные глыбки хроматина, ядрышко лежит эксцентрично, радиальная структура отсутствует. Не выделяет протеолитического фермента, питается бактериями, грибками, остатками растительной и животной пищи. В эндоплазме содержится много вакуолей. Эритроциты не заглатывает, даже если они содержатся в кишечнике в большом количестве (у больных бактериальной дизентерией). В нижнем отделе пищеварительного тракта образует восьми- и двухядерные цисты.
Ротовая амеба (Entamoeba gingival is).
Локализация. Ротовая полость, зубной налет у здоровых людей и имеющих заболевания полости рта, кариозные полости зубов.
Део графическое распространение. Повсеместно.
Морфофизиологическая характеристика. Вегетативная форма имеет размеры от 10 до 30 мкм, сильно вакуолизиро- нанную цитоплазму. Тип передвижения и строение ядра напоминают дизентерийную амебу. Эритроциты не заглатывает, питается бактериями, грибками. Кроме того, в вакуолях обнаруживают ядра лейкоцитов или так называемые слюнные тельца, которые после окраски могут напоминать эритроциты. Считают, что цист не образует. Патогенное действие в настоящее время отрицается. Обнаруживается в зубном налете здоровых людей в 60—70%. У людей с заболеваниями зубов и полости рта встречается чаще.
16.2. КЛАСС ЖГУТИКОВЫЕ (FLAGELLATA)
К жгутиковым относится наибольшее число патогенных для человека форм.
Морфофизиологическая характеристика. Имеют микроскопические размеры. Тело овальное, шаровидное или веретеновидное, покрыто, кроме наружной мембраны, тонкой оболочкой — пелликулой и сохраняет постоянную форму. Органоиды.движения — жгутики (1,2,4, 8 и более) — тонкие длинные выросты цитоплазмы, которые обычно начинаются на переднем конце тела. Жгутик состоит из свободной части, выходящей за пределы тела простейшего, и отдела, погруженного в эктоплазму, — базальное тело или кинетосома цилиндрической формы. У некоторых жгутиковых (лейшмании, трипаносомы) у основания жгутика, кроме того, помещается особый органоид — кинетопласт. По своей ульт£>аструкту- ре он соответствует митохондрии, но отличается высоким содержанием ДНК. Считают, что в кинетопласте происходит генерация энергии для движения жгутика, который совершает вращательное движение и как бы ввинчивается в воду. У некоторых представителей класса жгутик проходит вдоль тела, соединяясь с ним тонким выростом цитоплазмы. Упомянутый вырост, или ундулирующая мембрана, совершает волнообразные движения и служит дополнительным, органоидом движения.
По способу ассимиляции делятся на гетеротрофных, аутотрофных и миксотрофных. Гетеротрофные имеют пищеварительные вакуоли. Паразитические формы всасывают пищу всей поверхностью тела, в основном путем пиноцитоза. У аутотрофных органоидами питания служат хлоропласты, а миксотрофные могут использовать для питания как неорганические, так и органические вещества. Органоид выделения и осморегуляции у пресноводных свободноживущих — сократительная вакуоль, у паразитических форм она отсутствует. Обычно жгутиковые имеют по одному ядру, но встречаются двухядерные формы.
Размножение. Обычно бесполое, путем продольного деления на две части. У некоторых видов встречается половое размножение.
Обитают в пресной и морской воде. Многие формы являются паразитами человека и животных.
16.2.1. Отряд первичномонадные (Protomonadina) Род Лейшмания (Leishmania)
Наибольшее значение имеют представители рода Leishmania, относящегося к семейству Трипаносомвых.
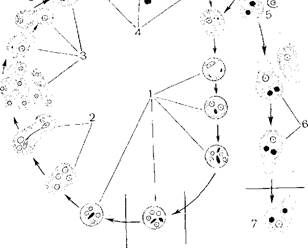
Отличительным признаком семейства Трипаносомовых служит способность образовывать в процессе цикла развития несколько морфологически различных форм в зависимости от условий существования. Смена форм происходит как в беспозвоночном, так и в позвоночном хозяевах.
Выделяют следующие морфологические формы: трипаносомную, критидиальную, лептомонадную, лейшманиальную и метациклическую (рис. 174).
Трипаносомная форма характеризуется сплющенным лентовидовым телом, в центре которого расположено овальное ядро. Жгутик начинается позади ядра. Осевая нить жгутика идет к переднему концу тела, образуя хорошо развитую ундулирующую мембрану. На переднем конце тела она заканчивается, а жгутик выступает вперед, образуя длинный свободный конец.
У критидиальной формы жгутик начинается немного впереди от ядра, направляясь вперед, образует короткую ундулирующую мембрану и свободный конец.
У лептомонадной формы жгутик начинается на самом краю переднего конца тела, ундулирующая мембрана отсутствует, свободный конец жгутика имеет значительную длину.
Лейшманиальная форма имеет округлую форму и крупное круглое ядро. Кинетопласт палочковидной формы расположен на переднем конце тела. Жгутик или отсутствует, или есть только его внутриклеточная часть, за пределы тела он не выходит.
Метацикличесжая форма сходна с критидиальной, но лишена свободного жгутика.
Жгутиконосцы рода Лейшманий имеют две морфологические формы — лептомонадную и лейшманиальную (рис. 175) или внутриклеточную.
Жизненный цикл проходит со сменой хозяев. Лейшманиальная форма паразитирует у человека и позвоночных, лептомонадная — в насекомых-переносчиках. Заболевания, вызываемые лейшманиями — лейшманиозы, являются природно-очаговыми, трансмиссивными.
Лейшмании подразделяются на дерматотропные (локализуются в коже) и висцеротропные виды (локализуются во внутренних органах).
Рис. 174. Формы жизненного цикла I рипаносомовых.
Внутри клеточная форма
Внеклеточные формы
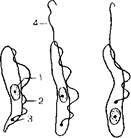
Рис. 175. Жгутиковые — паразиты человека.
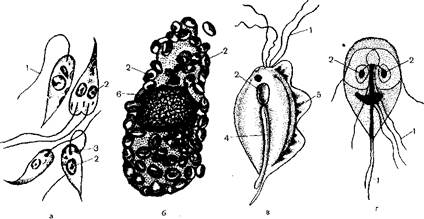
- лейшмания (лептомонадная форма); б — лсйшмания (лейшманиальная форма); в - фихомонада; г лямблия; I жгутик; 2 - ядро; 3 — кинетопласт; 4 — аксоетиль; 5 - ундулирующая мембрана; 6 — ядро тканевой клетки, пораженной лейишаниями.
[ - метациклическая (инвазионная); П — | ринаносомная; III — критидиальная; IV - лептомонадная; V - лейшманиальная; 1 — ядро; 2 — ундулирующая мембрана; 3 — ки- истопласт; 4 - свободный конец жгутика; Г) - внутриклеточный отдел жгутика; б — палочковидный кинетопласт.
Возбудитель висцерального лейшманиоза (Leischmania donovani).
Локализация. Клетки печени, селезенки, костного мозга, лимфатических узлов, ретикулоэндотелиальные клетки подкожной клетчатки.
Географическое распространен<