Этапы метаболизма
Условно процесс обмена веществ можно разделить на три этапа:
v Первый этап — ферментативное расщепление белков, жиров и углеводов до растворимых в воде аминокислот, моно- и дисахаридов, глицерина, жирных кислот и других соединений, происходящее в различных отделах желудочно-кишечного тракта, и всасывание их в кровь и лимфу.
Второй этап — транспорт питательных веществ кровью к тканям и клеточный метаболизм, результатом которого является их ферментативное расщепление до конечных продуктов. Часть этих продуктов используется для построения составных частей мембран, цитоплазмы, для синтеза биологически активных веществ и воспроизведения клеток и тканей. Расщепление веществ сопровождается выделением энергии, которая используется для процесса синтеза и обеспечения работы каждого органа и организма в
целом.
v Третий этап — выведение конечных продуктов метаболизма в составе мочи, кала, пота, через легкие в виде CO2 и т. д.
Пластический обмен - это совокупность реакций биосинтеза, или создание сложных молекул из простых. В клетке постоянно синтезируются белки из аминокислот, жиры из глицерина и жирных кислот, углеводы из моносахаридов, нуклеотиды из азотистых оснований и сахаров. Эти реакции идут с затратами энергии. Используемая энергия освобождается в ходе энергитического обмена. Энергетический обмен - это совокупность реакций расщепления сложных органических соединений до более простых молекул. Часть энергии, высвобождаемой при этом, идет на синтез богатых энергетическими связями молекул АТФ (аденозин-трифосфорной кислоты). Расщепление органических веществ осуществляется в цитоплазме и митохондриях с участием кислорода. Реакции ассимиляции и диссимиляции тесно связаны между собой и внешней средой. Из внешней среды организм получает питательные вещества. Во внешнюю среду выделяются отработанные вещества.
Процесс распада сложных веществ на более простые называется катаболизмом. Так, поступающие в пищей белки, жиры, углеводы под действием ферментов пищеварительного тракта распадаются на более простые составные части (аминокислоты, жирные кислоты и моносахариды). При этом высвобождается энергия. Обратный процесс, т. е. синтез сложных соединений из более простых называется анаболизмом. Он идет с затратой энергии. Из образовавшихся в результате пищеварения аминокислот, жирных кислот и моносахаридов в клетках синтезируются новые клеточные белки, фосфолипиды мембран и полисахариды.
Этапы энергетического обмена
1. Подготовительный - Под действием ферментов полисахариды расщепляются на моносахариды (глюкоза, фруктоза и Др.), жиры расщепляются до глицерина и жирных кислот, белки - до аминокислот, нуклеиновые кислоты до нуклеотидов. При этом выделяется небольшое количество энергии, которое рассеивается в виде тепла.
Бескислородный (анаэробное дыхание или гликолиз) — многоступенчатое расщепление глюкозы без участия кислорода. Его называют брожением. В мышцах в результате анаэробного дыхания молекула глюкозы распадается на две молекулы лировиноградной кислоты (С3Н4О3), которые затем восстанавливаются в молочную кислоту (С3Н6О3). В реакциях расщепления глюкозы участвуют фосфорная кислота и АДФ.
Суммарное уравнение этого этапа:
С6Н12О6 + 2Н3РО4 + 2АDФ →2С3Н6О3 + 2АТФ + 2Н2О
У дрожжевых грибков молекула глюкозы без участия кислорода превращается в этиловый спирт и диоксид углерода (спиртовое брожение). У других микроорганизмов гликолиз может завершаться образованием ацетона, уксусной кислоты и др. При распаде одной молекулы глюкозы образуется две молекулы АТФ, в связях которой сохраняется 40% энергии, остальная энергия рассеивается в виде тепла.
Энергетический обмен в аэробных клетках растений, грибов и животных протекает одинаково. Это свидетельствует об их родстве. Количество митохондрий в клетках тканей различно, оно зависит от функциональной активности клеток. Например, много митохондрий в клетках мышц.
Основной источник энергии в клетке - окисление субстратов кислородом воздуха. Этот процесс осуществляется тремя путями: присоединением кислорода к атому углерода, отщеплением водорода или потерей электрона. В клетках окисление протекает в форме последовательного переноса водорода и электронов от субстрата к кислороду. Кислород играет в этом случае роль восстанавливающегося соединения (окислителя). Окислительные реакции протекают с высвобождением энергии. Для биологических реакций характерны сравнительно небольшие изменения энергии. Это достигается за счет дробления процесса окисления на ряд промежуточных стадий, что позволяет запасать ее небольшими порциями в виде макроэргических соединений (АТФ). Восстановление атома кислорода при взаимодействии с парой протонов и электронов приводит к образованию молекулы воды.
Тканевое дыхание
Это процесс потребление клетками тканей организма кислорода, который участвует в биологическом окислении. Такой вид окисления называют аэробным окислением. Если конечным акцептором в цепи переноса водорода выступает не кислород, а другие вещества (например пировиноградная кислота), то такой тип окисления называют анаэробным.
Таким образом, биологическое окисление - это дегидрирование субстрата с помощью промежуточных переносчиков водорода и его конечного акцептора.
Дыхательная цепь - это переносчики протонов и электронов от окисляемого субстрата на кислород. Окислитель - это соединение, способное принимать электроны. Такая способность количественно характеризуется окислительно-восстановительным потенциалом по отношению к стандартному водородному электроду, рН которого равен 7,0. Чем меньше потенциал соединения, тем сильнее его восстанавливающие свойства и наоборот.
Т. о. любое соединение может отдавать электроны только соединению с более высоким окислительно-восстановительным потенциалом. В дыхательной цепи каждое последующее звено имеет более высокий потенциал, чем предыдущее.
Дыхательная цепь состоит из:
1. НАД - зависимой дегидрогеназы;
2. ФАД- зависимой дегидрогеназы;
3. Убихинона (КоQ);
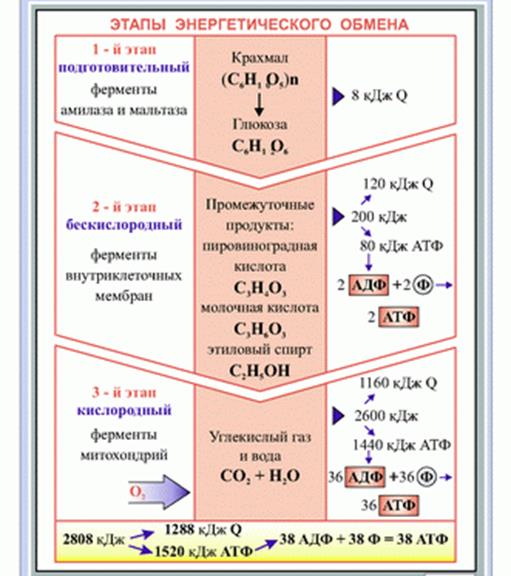
В этих реакциях, как и в реакциях бескислородного процесса, принимает участие Н3РО4 и АДФ и что кислородный процесс, как и бескислородный, сопряжен с синтезом АТФ. В ходе кислородного расщепления двух трехуглеродных молекул происходит образование 36 молекул АТФ - 36 богатых энергией фосфатных связей. Таким образом, суммарное уравнение кислородного процесса можно записать так:
2С3Н6О3+6О2+36Н3РО4+36АДФ =6СО2+6Н2О+36АТФ+36Н2О, а суммарное уравнение полного расщепления глюкозы так:
С6Н12О6+6О2+38Н3РО4+38АДФ =6СО2+6Н2О+38АТФ+38Н2О
Дыхание и горение. Окисление органических веществ, происходящее в клетке, часто сравнивают с горением: в обоих случаях происходит поглощение кислорода и выделение СО2 и Н2О. Однако между этими процессами имеются глубокие различия. Дыхание представляет высокоупорядоченный, многоэтапный процесс. Благодаря участию в нем ферментов оно идет с достаточной скоростью при температуре, несравненно более низкой, чем горение. Принципиально отличается в обоих процессах способ преобразования химической энергии расщепляемых веществ. При горении вся энергия переходит в тепловую. Дальнейшее использование ее для производства работы всегда происходит с низким к. п. д. При биологическом окислении главная часть энергии переходит в химическую энергию универсального энергетического вещества - АТФ, которое в дальнейшем используется клеткой с к. п. д., недостижимым для тепловых двигателей.
Световая фаза фотосинтеза происходит только на свету в мембране гран при участии хлорофилла, белков-переносчиков и АТФ-синтетазы. Под действием кванта света хлорофилл теряет электрон, переходя в возбужденное состояние.
Эти электроны передаются переносчиками (специальными молекулами) на наружную, т.е. обращенную к строме, поверхность мембраны тилакоидов, где накапливаются. Одновременно внутри полостей тилакоидов происходит фотолиз, т.е. разложение воды под действием энергии света:.
Ионы гидроксила отдают свои электроны, превращаясь в реакционноспособные радикалы ОН: Образующиеся электроны передаются переносчиками к молекулам хлорофилла и восстанавливают их, а радикалы ОН объединяются, образуя воду и свободный кислород:
Протоны водорода, образовавшиеся при фотолизе воды, не могут проникнуть через мембрану граны и накапливаются внутри нее, создавая и пополняя Н+-резервуар. В результате внутренняя поверхность мембраны граны заряжается положительно
(за счет Н+), а наружная — отрицательно (за счет е-). По мере накопления по обе стороны мембраны противоположно заряженных частиц увеличивается разность потенциалов между «обкладками» этого конденсатора. При достижении критической величины разности потенциалов сила электрического поля начинает проталкивать протоны через канал АТФ-синтетазы. На выходе из протонного канала создается высокий уровень энергии, которая используется для фосфорилирования имеющихся в строме пластид молекул АДФ:
Ионы водорода, оказавшись на наружной поверхности мембраны тилакоида, встречаются там с электронами, образуя атомарный водород, который идет на восстановление специфического переносчика НАДО (никотинамидадениндинуклеотидфосфата):Таким образом, во время световой фазы фотосинтеза происходят три процесса:
образование кислорода вследствие разложения воды,
синтез АТФ и
образование атомов водорода в форме НАДФ • Н2.
Кислород диффундирует в атмосферу, а АТФ и НАДФ • Н2 транспортируются в строму пластид и участвуют в процессах темновой фазы.
Темновая фаза – не совсем удачное название. Если мы говорим, что световая фаза названа так, чтобы подчеркнуть зависимость всех протекающих в ней реакций от света, то название «темновая фаза» означает, что все реакции, в ней протекающие, от света не зависят и проходят в темноте.
Но это не совсем точно. Очень многие реакции темновой фазы фотосинтеза зависят от света, так как ферменты, катализирующие эти реакции, индуцируются светом. Поэтому эту фазу фотосинтеза лучше называть путем превращения углерода, или циклом фиксации углерода, – по основному процессу, который в ней происходит.
(Здесь нужно обязательно пояснить, что происходит фиксация не чистого углерода, а углерода в составе углекислого газа СО2.)
Отметим, что разделение процесса фотосинтеза на две фазы происходит не только по отношению к свету, но и по месту протекания реакций. Реакции световой фазы протекают в тилакоидах гран и стромы, а реакции фиксации углерода протекают в матриксе (строме) хлоропластов.
Стоит обратить внимание учащихся на то, что в литературе встречается и другое название тилакоидов – ламеллы гран. Взаимосвязь терминов можно объяснить, зачитав отрывок из «Физиологии растений» Н.И. Якушкина: «Внутреннее пространство хлоропластов заполнено бесцветным содержимым – стромой – и пронизано мембранами (ламеллами). Ламеллы, соединенные друг с другом, образуют как бы пузырьки – тилакоиды. В хлоропластах тилакоиды двух типов. Короткие тилакоиды собраны в пачки и расположены друг над другом, напоминая стопку монет. Эти стопки называются гранами, а составляющие их ламеллы – ламеллами гран. Между гранами параллельно друг другу располагаются длинные тилакоиды. Составляющие их ламеллы получили название ламеллы стром».
Рассматривая Z-схему, мы установили, что конечные продукты циклического и нециклического фосфорилирования – АТФ и НАДФ . Н – используются в темновых реакциях фотосинтеза. Как же они используются?
Если в световой фазе АТФ и НАДФ . Н являются конечными продуктами, то в процессе фиксации углерода они используются на самом первом этапе всего цикла фиксации углерода. Весь цикл фиксации углерода можно представить в виде следующих стадий.
- Первая стадия – непосредственная фиксация углекислого газа – карбоксилирование.
- Вторая стадия – образование 3-фосфоглицеринового альдегида (ФГА).
- Третья стадия – образование продуктов фотосинтеза.
- Четвертая стадия – восстановление первоначальных реагентов.
Перечисленные стадии выделены условно – вместе они составляют цикл фиксации углерода, или цикл Кальвина.
В отличие от световых реакций, которые протекали в строгой последовательности, реакции фиксации углерода могут протекать параллельно, за исключением первых двух – фиксации углекислого газа и образования ФГА. Рассмотрим каждую стадию цикла.
Карбоксилирование
Эта стадия – ключевая, потому что в ней участвует СО2. Молекула углекислого газа соединяется с молекулой пятиуглеродного сахара рибулезодифосфата (РДФ) с образованием нестойкого шестиуглеродного соединения, которое затем распадается на две молекулы 3-фосфоглицериновой кислоты
Реакция карбоксилирования очень интересна тем, что в зависимости от условий она может протекать с образованием различных конечных продуктов. Так, например, при наличии СО 2 продуктом реакции будет только ФГК, а в присутствии О 2 РДФ не присоединяет углекислый газ и распадается на ФГК и фосфогликолевую кислоту, которая используется в процессах фотодыхания. Фотодыхание – это процесс, протекающий лишь на свету и сопровождающийся поглощением О 2 и выделением СО 2. Такое изменение хода реакции объясняется тем, что участвующий в ней фермент обладает двойной каталитической активностью – по отношению к углегислому газу и кислороду.
Этот фермент называется рибулозо-1,5-бифосфаткарбоксилаза-оксигеназа (РуБФ-карбоксилаза). Этот фермент составляет около 50% всех растворимых белков в листьях и потому может считаться самым распространенным белком в природе. Фермент состоит из двух субъединиц – большой и малой. Интересно, что белки больших субъединиц кодируются ДНК хлоропластов, а белки малых субъединиц – ядерной ДНК. Большие субъединцы обладают каталитической активностью и в отсутствие малых, которые, по-видимому, играют регуляторную роль. Этот факт может служить подтверждением того, что хлоропласты произошли от прокариотических предков.
Таким образом, на первых этапах фиксации углерода имеет место конкуренция между двумя процессами – фиксацией углерода и фотодыханием. Для сдвига баланса в сторону фиксации углерода необходимы ионы Мg 2 +