Секреторная теория. Берцелиус еще в 1890 г. считал почку железой, секретирующей воду и растворенные в ней вешества. У некоторых рыб отсутствует капиллярный клубочек, капилляры оканчиваются слепо.
Фильтрационная теория (К. Людвиг). Согласно фильтрационной теории, белки плазмы не могут проходить через почечный фильтр и остаются в кровотоке. Благодаря этому в плазме крови существует сила онкотического давления, равная 30 мм рт. ст., удерживающая воду в кровеносном русле и направленная против вектора гидростатического давления крови. Вторая сила, направленная против него же, обусловлена гидростатическим давлением ультрафильтрата — первичной мочи, заполняющей полость капсулы и почечных канальцев (20 мм рт. ст.).
Таким образом, эффективное фильтрационное давление образуется как результат алгебраической суммы всех составляющих:
рад - (ро„к + рП1.,р) = Рфи^гр. = 70 - (3° + 2°) = 20 мм рт. ст.
Теория активного транспорта. Объясняет образование ультрафильтрата за счет избирательной проницаемости и активного транспорта ингредиентов крови через трехслойный эпителий почечных капилляров и капсулы.
Скорость клубочковой фильтрации. Скорость клубочковой фильтрации определяется объемом ультрафильтрата, образующегося в почках за единицу времени. В среднем у мужчин скорость клубочковой фильтрации составляет 125 мл/мин, у женщин — 110 мл/мин.
Эффективность фильтрации поддерживается регуляцией почечного кровотока. Из каждого объема протекающей через почки плазмы крови 0,2 объема проходит через почечный фильтр и составляет клубочковый ультрафильтрат.
Первичная моча. В результате фильтрации за сутки образуется около 150 л первичной мочи, отличающейся от плазмы крови отсутствием белковых молекул, не проходящих через поры вели
чиной в 100 А. Первичная моча содержит необходимую организму воду и растворенные в ней вещества, большинство из которых представляют биологическую ценность, например: олигопептиды, аминокислоты, углеводы, соли и др. Лишь некоторые из веществ, растворенных в первичной моче, требуют выведения из организма. К ним относятся мочевина, мочевая кислота, креатинин, сульфаты и др.
Значение первичной мочи. В механизме фильтрации большого количества первичной мочи заложен определенный смысл.
• Удаление из крови вредных веществ, образующихся в результате метаболизма. Концентрация этих веществ в крови невелика, но при прохождении через почку больших объемов крови эти вещества выделяются, и кровь очищается от них.
• За счет обратного всасывания ингредиентов первичной мочи осуществляется тонкая регуляция задержки определенных солей и воды в организме или их удаление.
5.8.2.3. Канальцевая реабсорбция
Необходимые для организма биологически полезные вещества возвращаются (реабсорбируются) в кровь. В результате реабсорбции в сутки образуется 1,5—2,0 л конечной мочи, которая выделяется из организма. Остальной объем первичной мочи возвращается в кровоток.
Процесс реабсорбции начинается в проксимальных канальцах, куда поступает первичная моча из капсулы клубочка.
Порог выведения. Все вещества, содержащиеся в плазме крови, разделяются на пороговые и непороговые. К пороговым веществам относятся такие, которые выделяются в составе конечной мочи только при достижении определенной концентрации их в крови: например, глюкоза поступает в конечную мочу только в том случае, если ее содержание в крови превышает 6,9 ммоль/л.
Та концентрация вещества в крови, при которой оно не может быть полностью реабсорбировано и попадает в конечную мочу, получила название порог выведения.
Порог выведения для разных веществ различен и меняется при изменении функционального состояния организма.
Непороговые вещества в отличие от пороговых выводятся с мочой из организма при любой, даже самой низкой, концентрации
их в плазме крови. Примером таких веществ являются мочевина и креатинин.
Реабсорбция в проксимальных извитых канальцах. В этом отделе нефрона происходит обязательная, или облигатная, реабсорбция.
Процесс реабсорбции веществ из проксимальных канальцев в кровоток осуществляется за счет первичной реабсорбции натрия путем его активного транспорта. Активный транспорт натрия против концентрационного градиента связан с окислительными ферментативными процессами и протекает с участием фермента дегидрогеназы янтарной кислоты, который находится во всех клетках канальцевого эпителия.
Благодаря активному транспорту в кровь из первичной мочи всасываются также ионы К4, глюкоза и аминокислоты. Транспорт ионов Na+ на основе электростатического взаимодействия определяет также переход ионов О. Реабсорбция воды происходит пассивно вслед за натрием по осмотическому градиенту.
В проксимальных извитых канальцах реабсорбируется до 30 % натрия. Реабсорбируется 2/, объема ультрафильтрата. Объем мочи уменьшается в 8 раз, и одновременно увеличивается во столько же раз концентрация растворенных в ней веществ. В результате всасывания в кровоток воды повышается концентрация всех находящихся в моче веществ. Появляется концентрационный градиент между мочой, находящейся в канальцах, и плазмой крови, который обеспечивает движение растворенных в моче веществ в плазму крови за счет диффузии. Под влиянием концентрационного градиента пассивно за счет диффузии в кровь реабсорбируются аминокислоты, глюкоза, фосфаты, бикарбонаты и другие вещества. У человека скорость реабсорбции воды в проксимальных отделах нефрона составляет около 80% скорости клубочковой фильтрации.
В результате указанных процессов в проксимальных канальцах реабсорбируются многие аминокислоты, низкомолекулярные белки, витамины, большая часть ионов Na+, К+, Са2\ С1 и других веществ. Мочевина, мочевая кислота, аммиак реабсорбируются в очень малых количествах.
Процессы реабсорбции воды и процессы реабсорбции веществ сбалансированы, поэтому, в конечном счете, выходящая из проксимальных канальцев моча изотонична плазме крови.
Реабсорбция в петле нефрона. Дальнейший процесс концентрирования и уменьшения объема мочи происходит в петле нефрона (рис. 5.64).
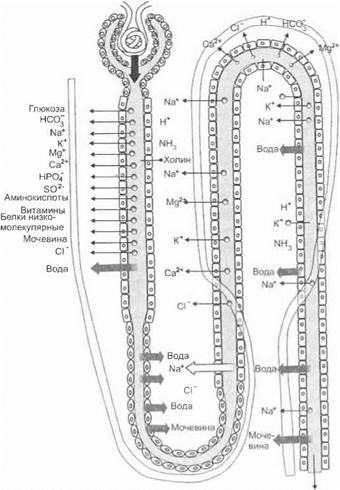
Рис. 5.64. Транспортные процессы (фильтрация, реабсорбция, секреция) в нефроне. Тонкими стрелками обозначены вещества, извлекаемые из крови канальцевым эпителием и секретируемые в просвет канальцев; толстыми стрелками — вещества, реабсорбируемые из первичной мочи в кровь
Концентрация первичной мочи в конечных отделах извитых проксимальных канальцев, как указывалось выше, изотонична крови почечной артерии. В нисходящем колене петли нефрона осмотическая концентрация мочи нарастает, достигая максимума в месте поворота петли: здесь она более чем в семь раз превышает осмотическую концентрацию крови почечной артерии. По мере продвижения мочи по восходящему колену петли нефрона в направлении от почечного сосочка к корковому слою осмотическая концентрация мочи вновь снижается. В месте перехода петли в дистальный извитой каналец она в три раза меньше осмотической концентрации крови.
Спускаясь затем по дистальному канальцу и особенно по собирательной трубочке к почечному сосочку, моча вновь приобретает высокую осмотическую концентрацию.
Осмотическая концентрация мочи повышается по мере ее продвижения от коркового слоя почек к сосочковому.
Поворотно-противоточная (умножительная) система почки. Принцип поворотно-противоточной системы известен в технике и используется, в частности, в теплообменниках.
Петля Генле работает как концентрирующий механизм. Благодаря своей конструкции петля обладает высокой способностью к концентрированию веществ при минимальной затрате энергии.
Два колена петли Генле — нисходящее и восходящее, — тесно соприкасаясь друг с другом, функционируют сопряженно, как один механизм. Эпителий нисходящего колена петли активно пропускает воду, но не ионы Na+. Эпителий же восходящего колена петли активно реабсорбирует ионы Na+, но не пропускает воду в тканевую жидкость. Вследствие реабсорбции воды из нисходящего колена петли Генле моча по ходу канала становится все более гипертоничной. Переход ионов Na+ из восходящего колена петли в тканевую жидкость, в свою очередь, повышает ее осмотическое давление и способствует тем самым еще более интенсивному выходу воды из нисходящего колена петли.
Таким образом, выход воды из мочи в тканевую жидкость в нисходящем колене петли Генле способствует реабсорбции натрия в восходящем колене, а реабсорбция натрия, в свою очередь, обусловливает выход воды в нисходящем колене.
Вследствие реабсорбции натрия в восходящем колене петли Генле, гипертоничная у вершины петли моча становится затем, в
конце восходящего колена петли Генле, изотоничной и даже гипо- тоничной по отношению к плазме крови.
В петле Генле моча, таким образом, теряет большое количество воды и натрия.
Реабсорбция в дистальных извитых канальцах и собирательных трубочках. В дистальных извитых канальцах нефрона происходит дальнейшее всасывание натрия, калия, воды, аминокислот, глюкозы и других веществ за счет тех же самых механизмов, что и при реабсорбции в извитых проксимальных канальцах. Реабсорбция здесь не постоянная, а зависит от уровня веществ в крови и в моче (факультативная реабсорбция).
В собирательных трубочках моча окончательно концентрируется благодаря пассивному току воды по осмотическому градиенту, создаваемому поворотно-противоточным механизмом (рис. 5.65).
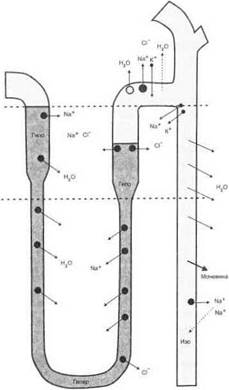 Рис. 5.65. Транспорт воды и ионов в петле Генле и собирательных трубочках нефрона
Рис. 5.65. Транспорт воды и ионов в петле Генле и собирательных трубочках нефрона
Реабсорбция глюкозы. Благодаря фильтрации глюкоза свободно проходит через почечный барьер и содержится в первичной моче в той же концентрации, что и в плазме крови. Однако в норме в конечной моче глюкозы нет или она содержится в ничтожных количествах. Из этого следует, что глюкоза полностью реабсорбируется в канальцах. В проксимальных извитых канальцах реабсорбируется до 98 % всей отфильтрованной глюкозы. Выделение глюкозы с мочой происходит тогда, когда содержание ее в крови превышает пороговую концентрацию, равную 1,8 г/л плазмы крови. При более высокой концентрации глюкозы в крови скорость выделения глюкозы с мочой прогрессивно возрастает.
Глюкоза поступает из просвета канальцев в клетки с помощью переносчика, который присоединяет ион Na+ по электрохимическому и концентрационному градиентам для Na+. Этому способствует работа натриево-калиевого насоса.
При превышении глюкозой пороговой величины содержания в крови и первичной моче она перестает реабсорбироваться. Этот порог обозначают как максимальный канальцевый транспорт (прежнее понятие — «почечный порог выведения»).
У мужчин максимальная скорость транспорта глюкозы в канальцах составляет в среднем 375 мг/мин, у женщин — 300 мг/мин.
Реабсорбция аминокислот. Механизмы реабсорбции аминокислот и глюкозы в общем однотипны. Аминокислоты свободно проходят через почечный фильтр. Реабсорбция аминокислот, так же как и реабсорбция глюкозы, сопряжена с переносом ионов Na+. Для разных групп аминокислот существуют различные транспортные молекулы, о чем свидетельствует конкурентное ингибирование реабсорбции одних аминокислот другими. В результате реабсорбции аминокислоты почти полностью реабсорбируются, и в конечной моче содержатся лишь их следы. Появление аминокислот в моче (аминоацидурия) может быть связано с нарушением деятельности транспортных механизмов.
Реабсорбция белков. Низкомолекулярные белки в небольших количествах (10—100 мг в 1 л фильтрата) в нормальных условиях могут проходить через почечный барьер. Однако в конечной моче белков в норме практически не содержится.
Механизм реабсорбции белков связан со способностью клеток проксимальных канальцев захватывать белки путем пиноцитоза, а
затем расщеплять их при участии лизосом. Таким путем может реабсорбироваться до 30 мг белков в 1 мин. При повреждении мембран клубочка и капсулы фильтрация белков возрастает и белок появляется в конечной моче (протеинурия).
Реабсорбция мочевины. Мочевина беспрепятственно фильтруется в клубочках. В проксимальных отделах нефрона в результате реабсорбции воды происходит концентрирование мочевины в моче. Благодаря трансканальцевому градиенту концентрации мочевина свободно поступает в кровь через клеточные мембраны путей пассивного транспорта в виде диффузии.
При существующей скорости движения фильтрата в канальцах общий объем реабсорбируемой мочевины сравнительно небольшой и недостаточен для выравнивания концентрации ее в плазме и фильтрате. Поэтому в физиологических условиях около 50 % профильтрованной мочевины удаляется с мочой. При замедлении или прекращении оттока мочи возрастает возврат мочевины в кровь. Удаление мочевины из организма зависит от объема выделенной мочи за определенное время.
В почках происходит внутрипочечный кругооборот мочевины. В собирательных трубочках под влиянием вазопрессина происходит реабсорбция воды, и концентрация мочевины повышается. Мочевина переходит в мозговое вещество почки. Затем по концентрационному градиенту мочевина диффундирует в петлю Генле и вновь поступает в дистальные канальцы и собирательные трубочки. При снижении всасывания воды в дистальных отделах нефрона мочевины выводится больше.
Реабсорбция воды, солей, кислот и оснований. Вода реабсорбируется во всех отделах нефрона. При нормальном употреблении воды с мочой выделяется не более 1 % объема фильтрата, образующегося за единицу времени, а 99 % реабсорбируется в кровоток. При избыточном потреблении воды диурез возрастает и может достигать 15% объема фильтрации, а канальцевая реабсорбция воды снижается до 85 %.
Ионы Na+, К+, Са2+, Mg2+ и НСО3_ содержатся в клубочковом фильтрате в тех же концентрациях, что и в плазме крови. В конечной моче остается очень небольшая часть этих ионов, подлежащих удалению. Большая часть их реабсорбируется, главным образом, в проксимальных извитых канальцах.
Реабсорбция слабых кислот и оснований зависит от их ионизации. Степень ионизации оснований увеличивается в кислой среде, и они с большей скоростью выделяются с кислой мочой. Слабые кислоты быстрее выводятся с щелочной мочой.
5.8.2.4. Секреторная функция канальцев
В процессе образования мочи ряд веществ (например, некоторые органические кислоты) не проникают в фильтрат, но, тем не менее, оказываются в конечной моче в результате канальцевой секреции.
В результате секреции в мочу поступают вещества из крови капилляров, окружающих канальцы или образующиеся в клетках канальцев (ионы водорода и аммиак). Секреция осуществляется за счет их активного транспорта канальцевым эпителием. Процесс сопряжен с биоэнергетическими ферментативными реакциями, направленными на образование АТФ. При угнетении тканевого дыхания секреция прекращается.
Известны три транспортные системы, действующие в проксимальных отделах нефронов, активно секретирующие различные (преимущественно инородные) вещества из крови. Одна из них осуществляет секрецию органических кислот (например, параа- миногиппуровой кислоты), йодсодержащих рентгеноконтрастных веществ (диодраст), пенициллина, красителей фенолового красного и др. Вторая — обеспечивает секрецию сложных органических оснований (тетраэтиламмония, N-метилникотинамида и др.), третья — секрецию этилендиаминтетраацетата (ЭДТА).
В целом состав образующейся в почках мочи определяется тремя процессами: клубочковой фильтрацией, канальцевой реабсорбцией и секрецией.
Процессы фильтрации, реабсорбции и секреции веществ, происходящие в почечных нефронах, представлены на рис. 5.64.
Моча по мере прохождения по канальцам почек приобретает кислую реакцию (pH = 4,5—6,5) вследствие того, что поры канальцевой реабсорбции двууглекислого натрия значительно шире, чем фосфатов. Реакция мочи может изменяться в зависимости от качества потребляемой пищи. При питании щелочной растительной пищей реакция мочи сдвигается в щелочную сторону. При питании мясной пищей или при интенсивной мышечной работе, наоборот, наблюдается возрастание кислотности мочи за счет поступления в
кровь увеличенных количеств молочных и фосфорных кислот, образующихся в мышцах.
Почки избирательно очищают плазму крови от некоторых веществ, концентрируя их в моче, которая удаляется из организма через мочевыводящие пути.
Конечная моча характеризуется следующими признаками:
• ряд веществ, входящих в состав плазмы крови, полностью отсутствует в конечной моче; в норме это белки, аминокислоты, глюкоза;
• некоторые вещества присутствуют в конечной моче в значительно больших концентрациях, чем в плазме: мочевина — в 65 раз, сульфаты — в 80 раз, мочевая кислота — в 12 раз; таким образом проявляется концентрирующая функция почек;
• состав и реакция мочи непостоянны: например, при алкалозе моча становится более основной, а при ацидозе — более кислой.
Для поддержания в крови постоянства концентрации водородных ионов и нейтрализации кислых продуктов обмена большое значение имеет синтез почкой аммиака. Аммиак связывает выделяемые с мочой кислотные радикалы, замещая натрий и калий, и образует аммонийные соли нелетучих кислот.
Фильтрационно-реабсорбционно-секреторная теория моче- образования. Согласно этой теории, содержание в конечной моче какого-либо вещества и выделение его складываются из объема клубочковой фильтрации и канальцевой секреции за вычетом его канальцевой реабсорбции.
По отношению к различным веществам возможны только фильтрация (например, для инулина), фильтрация с последующей реабсорбцией (например, для глюкозы), фильтрация с секрецией и сочетание всех процессов (например, ионы К+).
5.8.2.5. Методы функциональной оценки деятельности почек
Разработаны и широко используются методы количественной оценки функций почек.
Расчет коэффициента очищения. В процессе образования и выведения мочи из крови удаляется ряд веществ. Почечный клиренс отражает объемную скорость очищения определенного объема плазмы от того или иного вещества.
Этот показатель измеряется в мм за 1 мин и равен условному количеству плазмы крови, которая полностью очистилась отданного вещества за 1 мин.
Если вещество фильтруется и в дальнейшем не реабсорбируется и не секретируется, клиренс равен объему почечной фильтрации.
Если вещество после фильтрации реабсорбируется, клиренс будет меньше и равняется нулю в том случае, если вещество при реабсорбции полностью возвратилось в кровь. Если же вещество дополнительно секретируется и не реабсорбируется, клиренс увеличивается.
Рассмотрим пример расчета клиренса для мочевины. Концентрация мочевины в плазме крови и первичной моче равна 300 мг/л. Из каждого литра плазмы крови почка удаляет с мочой 18 мг мочевины. При указанной концентрации мочевины в плазме 18 мг ее содержится в 60 мл крови. Эта величина условно очищенной от мочевины крови и составляет клиренс по мочевине.
Расчет скорости клубочковой фильтрации. Скорость клубочковой фильтрации измеряется объемом фильтрата, образующегося в почках за единицу времени.
Для определения фильтрации используют диагностическое вещество инулин (полисахарид фруктозы), который вводят в кровоток. Инулин попадает в мочу только путем клубочковой фильтрации, не реабсорбируется и не секретируется, а также не претерпевает метаболических превращений в канальцах почек. Инулин беспрепятственно проходит почечный фильтр, не адсорбируется белками и содержится в фильтрате в той же концентрации, что и в плазме крови.
Количество инулина, профильтровывающееся за единицу времени, равно количеству этого вещества, удаленного с мочой:
С V = С х F
F — (С„н/1,х V„)/ C„H/wr,
где F— объем фильтрата за 1 мин; C„„/u — концентрация инулина в конечной моче; Уи — объем конечной мочи за 1 мин.
Расчет скорости канальцевой реабсорбции. Скорость канальцевой реабсорбции определяется количеством того или иного вещества, переносимого через стенку канальца в единицу времени. Она вычисляется с учетом разности между скоростью фильтрации вещества и скоростью его выделения с мочой. Для вычисления реабсорбции необходимо предварительно определить и знать объем фильтрации по инулину.
Скорость реабсорбции определяется для каждого вещества отдельно. Например, для глюкозы она равна:
C,,1/MxF = R + Crl/slVs„
где R — скорость реабсорбции в 1 мин.
R=CM/,1F-Cra/MxV>„
где Сгл/лл — концентрация глюкозы в плазме; F— объем почечной фильтрации за 1 мин по инулину; Сгл/и — концентрация глюкозы в моче; Им — диурез за 1 мин.
Из уравнения видно, что если реабсорбция равна фильтрации, то вещество полностью возвращается в кровь и в конечной моче оно отсутствует. В другом случае, если все профильтровавшееся вещество окажется в конечной.моче, реабсорбция будет равна нулю.
Расчет скорости секреции. При расчете скорости почечной секреции также используют данные об объеме фильтрации по инулину и исходят из того, что находящееся в конечной моче то или иное вещество может поступить только в результате фильтрации или секреции:
C,aF + S = C„xV.
Отсюда S (секреция вещества в 1 мин) равна:
S = C„xV4-CnlF,
где F— объем фильтрации по инулину; Сп,, — концентрация вещества в плазме; См — концентрация вещества в моче; — объем диффузии за 1 мин.
Расчет величины почечного кровотока. У взрослого человека массой тела 70 кг скорость кровотока в обеих почках составляет около 1300 мл/мин, что соответствует примерно 25 % общей объемной скорости кровотока в покое. Если учесть общую массу почек, равную 300 г, то видно, что скорость кровотока в почках значительно выше, чем в других крупных органах, таких как мозг, печень, сердце.
Интенсивный почечный кровоток необходим для обеспечения достаточно большого объема клубочковой фильтрации и не связан с метаболическими потребностями почек.
Определение почечного кровотока и плазмотока осуществляют с помощью веществ, вводимых в кровь, которые полностью удаляются из плазмы в мочу при однократном прохождении крови через почки.
Такое свойство наиболее выражено у парааминогиппуровой кислоты (ПАТ) и йодсодержащего рентгеноконтрастного вещества диодраста.
Количество поступающей из крови ПАГ равно ее концентрации в крови (Скр), умноженной на объем плазмотока (Еп1) в 1 мин.
Количество выделенной из почек ПАГ равно ее концентрации в моче (С„), умноженной на объем диуреза за 1 мин (Им).
С xV = С xV.
кр пл м м
Отсюда:
V„ = (CMxVM)/CKp.
При определении почечного кровотока вносят коррективы с учетом гематокрита (Г):
vKp = v,n/(i-r).
5.8.2.6. Саморегуляция почечного кровотока
Почечный кровоток обладает особой стабильностью. При изменении артериального давления от 80 до 180 мм рт. ст. почечный кро-
воток практически не меняется и начинает возрастать, только когда артериальное давление превышает 200 мм рт. ст.
Стабильность почечного кровотока при столь большом перепаде артериального давления определяется компенсаторными изменениями гемодинамического сопротивления почечных сосудов мышечного типа, которые сокращаются, расслабляются и сохраняют кровоток на постоянном уровне.
Саморегуляция почечного кровотока обеспечивает поддержание постоянства скорости клубочковой фильтрации.
Механизмы саморегуляции действуют в корковом веществе почек и практически отсутствуют в мозговом слое, где кровоток изменяется в соответствии с изменением общего артериального давления. Поскольку удельный вес кровотока в мозговом веществе почек составляет только 10% от общего кровотока, то в целом его роль невелика.
5.8.2.7. Регуляция мочеобразования
Деятельность почек зависит от почечных и внепочечных факторов регуляции.
Почечные факторы, определяющие почечную фильтрацию:
• количество функционирующих клубочков и проницаемость их канальцев;
• диаметр приносящего и выносящего сосудов;
• давление фильтрата в капсуле.
Внепочечные факторы, влияющие на почечную фильтрацию:
• функциональное состояние кровообращения, количество циркулирующей крови, величина артериального давления и скорость кровотока;
• степень гидратации организма, осмотическое и онкотическое давление;
• выделение воды через другие выделительные органы (потовые железы, желудок и пр.).
Почечные факторы, влияющие на реабсорбцию:
• реабсорбционная способность почечного эпителия;
• содержание пороговых и непороговых веществ в первичной моче.
Внепочечные факторы, определяющие реабсорбцию:
• эндокринная регуляция деятельности почечного эпителия со стороны желез внутренней секреции;
• содержание пороговых и непороговых веществ в крови.
Нервная регуляция. Нервная регуляция деятельности почек осуществляется вегетативной нервной системой через блуждающие и чревные нервы.
Активация симпатической нервной системы вызывает сужение сосудов почки, уменьшение фильтрации и снижение диуреза. Увеличивается канальцевая реабсорбция натрия и воды. Денервация (перерезка чревного нерва) увеличивает диурез на стороне операции (опыт К. Бернара). В результате денервации устраняются сосудосуживающие влияния и угнетается реабсорбция натрия.
В отдельных случаях активация симпатической нервной системы может вызвать временное прекращение диуреза, например, на фоне психоэмоционального стресса.
Раздражение волокон блуждающих нервов вызывает усиление реабсорбции глюкозы и секреции органических кислот.
Перерезка блуждающих нервов приводит к увеличению содержания хлористого натрия в моче. Раздражение блуждающих нервов вызывает снижение содержания хлористого натрия в моче (Л. А. Ор- бели). Однако не совсем ясно, что при этом первично — сосудистые изменения или изменения механизма мочеобразования.
Болевые реакции, как правило, прекращают мочеобразование. Показана возможность выработки условного рефлекса усиления диуреза при введении воды в желудок.
Гуморальная регуляция. Гуморальная регуляция деятельности почек в процессах мочеобразования является основной.
Влияние антидиуретического гормона — вазопрессина. Одним из важнейших гормонов, влияющих на функции почек, является антидиуретический гормон (АДГ) — вазопрессин. Действуя на V,-рецепторы собирательных трубочек, АДГ уменьшает диурез, сберегает воду в организме и повышает концентрацию мочи. АДГ повышает также реабсорбцию мочи в дистальных отделах нефронов за счет увеличения проницаемости для воды эпителия дистальных извитых канальцев и собирательных трубочек.
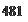 В отсутствие АДГ дистальные отделы нефрона почти непроницаемы для воды и реабсорбции воды практически не происходит.
В отсутствие АДГ дистальные отделы нефрона почти непроницаемы для воды и реабсорбции воды практически не происходит.
1 6 Нормальная физиология
При этом образуется большое количество гипотонической мочи. Максимальный диурез в этом случае может достичь 15% объема клубочковой фильтрации, т.е. 25 л/сут. Это наблюдается у больных несахарным мочеизнурением, характеризующимся недостаточностью АДГ. Таким образом, 15 % общей реабсорбции воды регулирует АДГ, а 85 % — сохраняется как обязательная реабсорбция даже при отсутствии АДГ.
Механизм действия АДГ заключается в стимуляции секреции гиалуронидазы клетками собирательных трубочек. Этот фермент деполимеризует гиалуроновые структуры соединительной ткани, увеличивая их проницаемость для воды и тем самым обеспечивая движение воды по осмотическому градиенту. Объем реабсорбции зависит от концентрации АДГ в крови.
Действие АДГ опосредовано цАМФ. Точкой приложения действия АДГ является аденилатциклаза — фермент, катализирующий превращение АТФ в цАМФ.
Влияние гормонов коры надпочечников. Деятельность почки находится под контролем минералокортикоидов и глюкокортикоидов коры надпочечников. Одним из наиболее эффективных минералокортикоидов является альдостерон. Этот гормон регулирует реабсорбцию натрия, секрецию калия и ионов водорода в канальцах почек. Под влиянием альдостерона в проксимальных канальцах происходит задержка натрия и воды в организме. Напротив, угнетение функции надпочечников, а в эксперименте — удаление надпочечников приводят к повышению выделения с мочой натрия и снижению экскреции калия. В результате возникает значительная потеря организмом натрия и воды. Компенсаторное введение альдостерона восстанавливает баланс этих электролитов.
Альдостерон также уменьшает реабсорбцию кальция и магния в проксимальных канальцах.
Механизм действия альдостерона заключается в его проникновении через мембраны в клетки и взаимодействии с ядерными белками-рецепторами. Действие альдостерона проявляется в стимуляции ДНК-зависимого синтеза информационной, транспортной и рибосомальной РНК. Последние участвуют в синтезе на рибосомах нового белка, который обеспечивает активность натриевого насоса, осуществляющего транспорт ионов Na+.
При введении в организм веществ, блокирующих синтез белка (например, пуромицина), эффекты альдостерона исчезают.
Гормоны коры надпочечников способствуют увеличению диуреза, усиливая фильтрацию и уменьшая реабсорбцию.
Кортикостероиды оказывают стимулирующее влияние на мо- чеобразование.
Паратгормон увеличивает реабсорбцию кальция и тормозит реабсорбцию фосфатов в почечных канальцах. Паратгормон угнетает реабсорбцию ионов Na+ и НСО3 в проксимальных канальцах и активирует реабсорбцию магния в восходящем колене петли Генле. Тиреокальцитонин способствует экскреции кальция и фосфата почками.
Натрийуретический гормон усиливает выделение ионов Na+ и воды с мочой.
5.8.2.8. Участие почек в различных функциональных системах
Почки участвуют в различных функциональных системах организма. С деятельностью почек связано поддержание ряда жизненно важных показателей организма: ионного состава и объема межклеточной жидкости, осмотического давления и водно-солевого баланса, pH, объема циркулирующей крови, величины артериального давления, содержания продуктов белкового обмена и концентрации в крови азотистых веществ, выведения чужеродных, токсичных веществ из организма (см. разд. 4.3., 5.2.12., 5.3.7).
В указанных функциональных системах почки выступают в качестве исполнительного звена саморегуляции. Можно полагать, что качественные особенности мочеобразования в разных функциональных системах различны. Вопрос этот требует специальных исследований.
Инкреторная функция почек. В эпителиальных клетках проксимальных канальцев почки образуется витамин D3, стимулирующий в эпителиальных клетках кишечника образование кальцийс- вязывающих и кальцийтранспортных белков, необходимых для всасывания ионов Са2+. В перитубулярных клетках почек образуется эритропоэтин, стимулирующий образование эритроцитов.
Ренин-ангиотензиновая система. Юкстагломерулярные нефроны наряду с мочеобразованием выполняют эндокринную функцию. Юкстагломерулярными эпителиоидными клетками вырабатывается ренин, представляющий собой фермент протеазу, поступающий в кровоток. Под действием ренина от находящегося в плазме крови ангиотензиногена отщепляется ангиотензин I. Ангиотензин I под действием превращающего фермента, активность которого в крови и легких высока, переходит в активную форму — ангиотензин II, который затем расщепляется протеолитическими ферментами, в результате чего в крови появляется ангиотензин III и в последующем — неактивные фрагменты. Ангиотензин II представляет собой вещество, обладающее наиболее выраженным сосудосуживающим эффектом, обусловливающим значительное повышение артериального давления.
Наряду с этим ангиотензин II стимулирует выброс альдостерона корой надпочечников. Альдостерон, как указывалось выше, повышает реабсорбцию натрия в почечных канальцах и задерживает воду в организме. В результате этого также повышается системное артериальное давление.
Ренин-ангиотензиновая система активируется при падении артериального давления, недостатке солей в организме и появлении гиповолемии. Все эти состояния тесно связаны между собой.
Стимуляция выделения ренина юкстагломерулярными клетками осуществляется несколькими факторами:
• ишемизация почек, сужение почечной артерии являются причинами развития почечной гипертензии, при которой систолическое давление может достичь 200 мм рт. ст. и выше. В эксперименте почечную гипертензию можно вызвать путем искусственного стеноза, частичного пережатия почечных артерий (гипертензия Гольдблата);
• при увеличении артериального давления включается внутрипочечный барорецепторный механизм;
• выделение ренина увеличивается при снижении объема циркулирующей крови. Считают, что в этом процессе участвуют клетки плотного пятна юкстагломерулярного аппарата и их p-адренорецепторы;
• выработка ренина увеличивается при активации симпатических нервов и в эксперименте — внутриартериальном введении норадреналина и адреналина;
• различные гуморальные факторы, содержащиеся в протекающей через почки крови, также влияют на синтез и высвобождение ренина.
Интенсивная выработка ренина зависит от содержания ионов Na+ и К+ в плазме крови и клетках плотного пятна.
Наряду с факторами, активирующими выделение ренина, проявляются и тормозные влияния, в частности со стороны ангиотензина II и АДГ, которые по механизму отрицательной обратной связи ограничивают действие ренина.
Основная роль ренин-ангиотензиновой системы в целостной регуляции гемодинамических показателей заключается в повышении артериального давления, сохранении и задержке в организме натрия и увеличении объема внеклеточной жидкости.
Непосредственно в деятельности почек ренин обеспечивает процесс мочеобразования, как только возникает угроза снижения или прекращения диуреза из-за падения артериального давления.
Предсердный натрийуретический пептид. При повышении общего уровня артериального давления в предсердиях вырабатывается предсердный натрийуретический пептид, который поступает в кровь и усиливает диурез, вследствие задержки реабсорбции натрия и воды в канальцах почек. В результате общее кровяное давление снижается.
Натрийуретический пептид определяет следующие процессы в почках:
• оказывает тормозное действие на секрецию альдостерона, благодаря чему увеличивает содержание в конечной моче натрия;
• увеличивает скорость фильтрации в почечных клубочках, что приводит к увеличению объема конечной мочи;
• уменьшает секрецию ренина клетками юкстагломерулярного комплекса;
• увеличивает выделение с мочой хлоридов, магния и кальция.
Почечная недостаточность. Функциональные возможности почек имеют значительный резерв. Одна здоровая почка способна обеспечить нормальное существование человека. Появление почечной недостаточности развивается в том случае, если число функционирующих нефронов составляет менее 30 % по сравнению с нормальным числом, а скорость фильтрации — ниже 50 мл/мин. При почечной недостаточности происходят задержка в организме азотсодержащих веществ (моче