Содержание
Понятия экосистема и энергия ……………………………………………………2
Поступление энергии в экосистему, понятие биологическая продуктивность..4
Пищевые цепи………………………………………………………….…………...6
Поток энергии в экосистеме………………………………………………………..8
Круговорот веществ в экосистеме…………………………………………………13
Литература…………………………………………………………………………..20
Понятия экосистема и энергия
Прежде чем говорить об энергетике экосистем, необходимо рассмотреть такое основополагающие для данной темы понятия, как экосистема и энергия. Понятие экосистемы было введено в науку английским ученым А. Тенсли в 1935 году. Экосистема включает в себя все живые организмы, совместно функционирующие на какой-либо конкретной територии, а также из компонентов неживой природы, определяющих характер местноти, в которой расположена экосистема. Организменная часть экосистемы взаимодействует с ее неживыми сотавляющими, в результате чего под воздействием энергии, притекающей извне, происходит круговорот веществ между живой и неживой частями экосистемы и складывается ее внутренняя структура. Экосистема — основная функциональная и структурная единица живой природы, носитель ее элементарных свойств. Экосистемы различаются между собой по своим размерам, самой крупной экосистемой является биосфера.
В любой экосистеме седует выделить несколько составляющих ее компонентов. Первый из них — неорганические вещества (углерод, кислород, азот, углекислый газ, вода и т.д.). Затем следуют органические соединения (белки, липиды (жиры), углеводы и др.), являющиеся связующим звеном между живой (биотической) и неживой (абиотической) частями экосистемы. К важным ее элементам относятся и физические факторы абиотической среды (температура, влажность, давление и др.). Биотическая часть экосистемы состоит из живых организмов, подразделяющихся на три основные категории:
|
|
1) Продуценты — организмы, поддерживающие свою жизнедеятельность, путем производства из неорганических веществ органические при помощи химической реакции фотосинтеза, для осуществления которой необходим приток световой энергии.
2) Консументы — живые существа, поедающие другие оранизмы или частицы органичекого вещества и, таким образом, обеспечивающие себя необходимой энергией.
3) Редуценты — организмы, питающиеся останками растений и животных (т.е. органическим веществом) и разлагающие их до простых минеральных веществ, которые могут легко усваиваться продуцентами.
Первые три вышеназванных компонента представляют собой абиотическую составляющую экосистемы, последние же три являютя ее организменной частью. Причем продуценты называют автотрофами, так как они самостоятельно вырабатывают органическое вещесвто, а консументы и редуценты относятся к гетеротрофам, то есть к организмам, потребляющим готовое органическое вещество. Продуценты, консументы и редуценты образуют собой так называемую биомассу. Совокупность всех шести данных элементов складывают собой структуру экосистемы.
Одно из определений энергии характеризует ее, как общую количественную меру движения и взаимодействия всех видов материи, благодаря чему все явления природы связаны воедино. Изменение энергии какой — либо системы происходит при совершении работы. Иными словами, энергия — это способность совершать работу. Свойства энергии описываются первым и вторым законами термодинамики.
|
|
Первый закон термодинамики сводится к следующему утверждению: энергия может переходить из одной формы в другую, но она не исчезает и не создается заново. Данный закон называют также законом сохранения энергии. Этому закону подчиняются все известные процессы в природе.
Второй закон термодинамики гласит: поскольку некоторая часть энергии всегда рассеивается в виде недоступной для использования тепловой энергии, эффективность самопроизвольного превращения кинетической энергии в потенциальныю всегда меньше 100%. Количественная мера энергии, которая становится недоступной для использования, называется энтропией. Способность создавать и поддерживать высокую степень внутренней упорядоченности, то есть состояние с низкой энтропией — важнейшая характеристика экосистемы. Подводя итог вышесказанному, необходимо отметить, что экосистемы представляют собой открытые неравновесные термодинамические системы, постоянно обменивающиеся с внешней средой веществом и энергией. При этом, они уменьшают энтропию внутри себя, но увеличивают ее снаружи, в соответствии с законами термодинамики. Следовательно, энергетика экосистемы — это непрерывный процесс, в ходе которого энергия поступает в экосистему извне, подвергается преобразованию в ней и выделяется из нее.
Поступление энергии в экосистему, понятие биологическая продуктивность
|
|
Энергия поступает в экосистему в основном за счет солнечного излучения и длинноволнового теплового излучения от близлежащих тел. Солнечный свет падает на землю из космоса с энергией 1,98 ккал/см^2 в минуту. Но до земли доходит максимум 67% от данной энергии. Поступление солнечной энергии к экосистеме составляет в среднем 300-400 ккал/см^2 в день. Но в процессе фотосинтеза, обеспечивающем энергией живые компоненты экосистемы, усваивается лишь 1% всей поступившей к экосистеме энергии. Остальная часть энергии уходит на поддержание определенной температуры окружающей среды, приводит в действие системы погоды, круговорота воды и т.д.
Солнечная энергия, поступившая к экосистеме, усваивается категорией живых организмов, известных нам под называнием продуценты. Скорость, с которой продуценты поглощают лучистую энергию солнечного излучения, а, значит, и скорость, с которой в экосистеме происходит накопление органического вещества, определяют, как биологическую продуктивность. Первичную продуктивность P выражают в единицах массы, энергии или эквивалентных единицах в промежуток времени.
В процессе производства органического вещества в экосистеме выделяют четыре последовательных уровня:
1) Валовая первичная продуктивность — скорость накопления в процессе фотосинтеза органического вещества (или биомассы), с учетом той его части, которая за время измерений была израсходована на дыхание (то есть на поддержание жизнедеятельности биомассы)
2) Чистая первичная продуктивность — скорость накопления органического вещества в организмах продуцентов, за вычетом той его части, которая была израсходована ими на дыхание.
3) Вторичная продуктивность сообщества — скорость накопления органического вещества на уровне консументов, которые, как известно, питаются готовой биомассой.
4) Чистая продуктивность сообщества — скорость накопления органического вещества, не потребленного гетеротрофами. Эта величина равна разности между чистой первичной продуктивностью сообщества и количеством вещества, не потребленного гетеротрофами. Чистая продуктивность сообщества в определенный момент времени выражается наличной биомассой. Другое название чистой продуктивности сообщества — урожай на корню.
Продуктивность экосистемы зависти от ряда факторов. Высокие скорости продуцирования наблюдаются в экосистемах, где физические факторы благоприятны, особенно в случая поступления экосистеме дополнительной энергии извне. Поступление энергии со стороны абиотических компонентов экосистемы уменьшают затраты живых организмов на поддержание собственной жизнедеятельности. Для оценки продуктивности экосистемы необходимо также учитывать утечку энергии с урожаем продукции, а также стрессовые воздействия, уменьшающих долю энергии, задействованной в процессе производства биомассы.
Также необходимо рассмотреть баланс между валовой первичной продукцией и дыханием сообщества. Здесь нужно сделать допущение о том, что вся чистая первичная продукция потребляется консументами первого порядка(т.е. консументами, питающихся биомассой продуцентов). Делая аналогичные допущения для последующих уровней питания (так называемых трофических уровней), следует записать следующую систему уравнений:
Pпр = Pк1 + Rпр
Pк1 = Pк2 + Rк1
Pк2 = Pк3 + Rк2
…………
Pпр = Rпр + Rк1 + Rк2 +…+ Rкn
Суммирование равенств показывает, что вся валовая первичная продукция полностью расходуется на дыхание сообщества без остатка, то есть все произведенное в экосистеме органическое вещество было полностью потреблено. Данное равенство между производством биомассы и ее потреблением представляет собой иллюстрацию стабильного функционирования любой экосистемы. Примером подобной стабильности может служить экосистема тропического леса, где вся произведенная продуцентами биомасса полностью потребляется гетеротрофными организмами. Но, обычно, производство биомассы преобладает над ее потреблением, в результате чего происходит накопление непотребленного органического вещества (например, в форме каменного угля).
Цепи питания
Перенос энергии в экосистеме осуществляется через так называемые пищевые цепи. В свою очередь, пищевая цепь — это перенос энергии от ее первоначального источника (обычно им являются автотрофы) через ряд организмов, путем поедания одних другими. Пищевые цепи подразделяются на два вида:
1) Пастбищная пищевая цепь. Она начинается с растений и тянется дальше к растительноядным животным (фитофагам), а затем и к хищникам. В подобной цепи при каждом переходе к следующему звену теряется до 80-90% потенциальной энергии пищи, так как она переходит в тепло. Пастбищные пищевые цепи делятся на пищевые цепи хищников и пищевые цепи паразитов. При продвижении по пищевой цепи хищников, размер каждого последующего его члена больше, чем размер предыдущего, но численность каждых следующих участников пищевой цепи меньше численности ее предыдущих представителей. Примером пищевой цепи хищников может служить следующая последовательность:
Сосна обыкновенная => Тли => Божьи коровки => Пауки =>Насекомоядные
птицы => Хищные птицы.
В отличии от пищевой цепи хищников, пищевые цепи паразитов ведут к организмам, которые все более уменьшаются в размерах и увеличиваются численно. В качестве примера можно привести следующую цепь:
Трава => Травоядные млекопитающие => Блохи => Жгутиконосцы.
2) Детритная пищевая цепь. Она берет свое начало от мертвого органического вещества (т.н. детрита), которое либо потреблятеся в пищу мелкими, преймущественно беспозвоночными животными, либо разлагается бактериями или грибами. Организмы, потребляющие мертвое органическое вещество, называются детритофагами, разлагающие его — деструкторами.
Пастбищная и детритная пищевые цепи обычно существуют в экосистемах совместно, но один из видов пищевых цепей почти всегда доминирует над другим. В некоторых же специфических средах (например в подземной), где из-за отсутствия света невозможна жизнедеятельность зеленых растений, существуют только детритные пищевые цепи.
В экосистемах пещевые цепи не изолированы друг от друга, а тесно переплетены. Они составляют так называемые пищевые сети. Это происходит потому, что каждый продуцент имеет не одного, а нескольких консументов, которые, в свою очередь, могут иметь несколько источников питания. Взаимосвязи внутри пищевой сети наглядно иллюстрирует приведенная ниже схема.
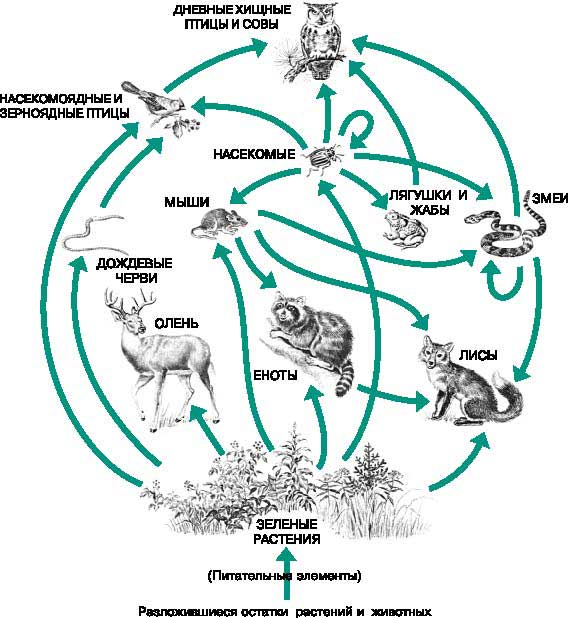
Схема пищевой сети.
В пищевых цепях образуются так называемые трофические уровни. Трофические уровни классифицируют организмы в пищевой цепи по типам их жизнедеятельности или по источникам получения энергии. Растения занимают первый трофический уровень (уровень продуцентов), травоядные (консументы первого порядка) относятся ко второму трофическому уровню, хищники, поедающие травоядных, образуют третий трофический уровень, вторичные хищники — четвертый и т.д. первого порядка.
Поток энергии в экосистеме
Как нам известно, перенос энергии в экосистеме осуществляется через пищевые цепи. Но далеко не вся энергия предыдущего трофического уровня переходит на следующий. В качестве примера можно привести следующую ситуацию: чистая первичная продукция в экосистеме (то есть количество энергии, накопленное продуцентами) составляет 200 ккал/м^2, вторичная продуктивность (энергия, накопленная консументами первого порядка) равна 20 ккал/м^2 или 10% от предыдущего трофческого уровня, энергия же следующего уровня составляет 2 ккал/м^2, что равно 20% от энергии предыдущего уровня. Как видно из данного примера, при каждом переходе на более высокий уровень теряется 80-90% энергии предыдущего звена пищевой цепи. Подобные потери связаны с тем, что значительная часть энергии при переходе с одной ступени на другую не усваивается представителями следующего трофического уровня или превращается в тепло, недоступное для использования живыми организмами.
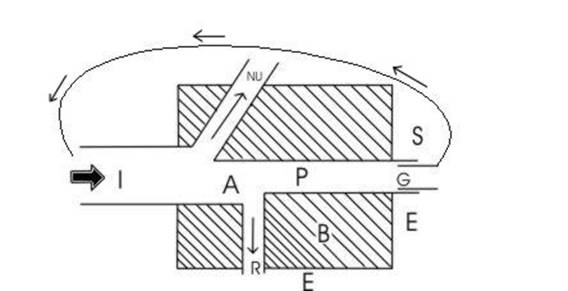
Универсальная модель потока энергии.
Поступление и расход энергии можно рассмотреть с помощью универсальной модели потока энергии. Она применима к любому живому компоненту экосистемы: растению, животному, микроорганизмам, популяции или трофической группе. Подобные графические модели, соединенные между собой, могут отражать пищевые цепи (при последовательном соединении схем потока энергии нескольких трофических уровней образуется схема потока энергии в пищевой цепи) или биоэнергетику в целом. Поступившая в биомассу энергия на схеме имеет обозначение I. Однако, часть поступившей энергии, не подвергается превращнию (на рисунке обозначена, как NU). Например, это происходит в случае, когда часть света, проходящего через растения, не поглощается ими, или когда часть пищи, проходящей через пищеварительный тракт животного, не усваивается его организмом. Усвоенная (или ассимилированная) энергия (обозначенная за A) используется для различных целей. Она тратитися на дыхание (на схеме- R) т.е. на поддержание жизнедеятельности биомассы и на продуцирование органического вещества (P). Продукция, в свою очередь, принимате различные формы. Она выражается в энергетических затратах на рост биомассы (G), в различных выделениях органического вещетсва во внешнюю среду (E), в запасе энергии организмом (S) (примером подобного запаса являются жировые накопления). Запасенная энергия образует на схеме так называемую рабочую петлю, так как данная часть продукции используется для обеспечения энергией в будущем (напимер, хищник использует свой запас энергии для поиска новых жертв). Оставшаяся часть продукции представляет собой биомассу (B).
Универсальную модель потока энергии можно интерпретировать двояко. Во-первых она может представлять популяцию какого-либо вида. В данном случае каналы потока энергии и связи рассматриваемого вида с другими видами представляют собой схему пищевой цепи. Другая интерпритация трактует модель потока энергии как изображение какого-либо энергетического уровня. Тогда прямоугольник биомассы и каналы потока энергии представляют все популяции, поддерживаемые одним и тем же источником энергии.
Для того, чтобы наглядно показать различие подходов трактовки универсальной модели потока энергии можно рассмотреть пример с популяцией лис. Часть рациона лисиц составляет растительность (плоды и т.д.), другую же часть составляют травоядные животные. Чтобы подчеркнуть аспект внутрипопуляционной энергетики (первая интерпритация энергетической модели), всю популяцию лис следует изобразить в виде одного прямоугольника, если же нужно распределить метаболизм (метаболизм — обмен веществ, интенсивность обмена веществ) популяции лис на два трофических уровня, то есть отобразить соотношение ролей растительной и животной пищи в обмене веществ, необходимо построить два или несколько прямоугольников.
Зная универвальную модель потока энергии, можно определить отношение величин энергетического потока в разных точках пищевой цепи.Выраженные в процентах, эти отношения называют экологической эффективностью. Существует несколько групп экологических эффективностей. Первая группа энергетических отношений: B/R и P/R. Доля энергии, расходущейся на дыхание, велика в популяциях крупных организмов. При стрессовом воздействии внешней среды R возрастает. Величина P значительна в активных популяциях мелких организмов (например водорослей), а также в системах, получающих энергию извне.
Следующая группа отношений: A/I и P/A. Первое из них называется эффективностью ассимиляции (т.е. эффективностью использования поступившей энергии), второе — эффективностью роста тканей. Эффективность ассимиляции может варьироваться от 10 до 50% и выше. Она может либо достигать малой величины (при ассимиляции энергии света растениями), либо иметь большие значения (при ассимиляции энергии пищи животными). Обычно эффективность ассимиляции у животных зависит от их пищи. У растительноядных животных она достигает 80% при поедании семян, 60% при использовании в пищу молодой листвы, 30-40% — более старых листьев, 10-20% при питании древесиной. У хищных животных эффективность ассимиляции составляет 60-90%, так как животоная пища гораздо легче усваивается организмом, чем растительная.
Эффективность роста тканей также широко варьируется. Наибольших значений она достигает в тех случаях, когда организмы имеют небольшие размеры и условия среды их обитания не требуют больших энергетических затрат на поддержание оптимальной для роста организмов температуры.
Третья группа энергетических отношений: P/B. Если рассматривать P как скорость прироста продукции, P/B представляет собой отношение продукции в конкретный момент времени к биомассе. Если расчитывается продукция за определенный промежуток времени, значение отношения P/B определяется исходя из средней за этот промежуток времени биомассы. В данном случае P/B является безразмерной величиной и показывает, во сколько раз продукция больше или меньше биомассы.
Следует отметить, что на энергетические характеристики экосистемы оказывает влияние размеры организмов, населяющих экосистему. Установлена зависимость между размером организма и его удельным метаболизмом (метаболизмом на 1г. биомассы). Чем мельче организм, тем выше его удельный метаболизм и, следовательно, тем меньше биомасса, которая может поддерживаться на данном трофическом уровне экосистемы. При одинаковом количестве использованной энергии организмы больших размеров накапливают большую биомассу, чем мелкие. Например, при равном значении потребленной энергии, биомасса, накопленная бактериями, будет гораздо ниже биомассы, накопленной крупными организмами (наприемр млекопитающими). Иная картина открывается при рассмотрении продуктивности. Так как продуктивность — это скорость прироста биомассы, то она больше у мелких жвотных, которые имеют более высокие темпы размножения и обновления биомассы.
В связи с потерей энергии внутри пищевых цепей и зависимостью метаболизма от размера особей, каждое биологическое сообщество приобретает определеную трофическую структуру, которая может служить характеристикой экосистемы. Трофическая структура характеризуется или урожаем на корню, или количеством энергии, фиксируемой на единицу площади в единицу времени каждым последующим трофическим уровнем. Трофическую структуру можно изобразить графически в виде пирамид, основанием у которых служит первый трофический уровень (уровень продуцентов), а последующие трофические уровни образуют "этажи" пирамиды. Выделяют три типа экологических пирамид.
1) Пирамида численности (на схеме обозначена цифрой 1) Она отображает количество отдельных организмов на каждом из трофических уровней. Численность особей на разных трофических уровнях зависит от двух основных факторов. Первый из них — более высокий уровень удельного метаболизма у мелких животных по сравнению с крупными, что позволяет им иметь численное превосходство над крупными видами и более высокие темпы размножения. Другой из вышеназванных факторов — существование у хищных животных верхнего и нижнего предела размера их жертв. Если жертва намного крупнее хищника по размерам, то он будет не в состоянии ее одолеть. Добыча же небольшого размера не сможет удовлетворить энергетических потребностей хищника. Поэтому для каждого хищного вида существует оптимальный размер жертв Однако, для данного правила существуют исключения (например, змеи с помощью яда убивают животных, превышающих их по размерам). Пирамиды чисел могут быть обращены "острием" вниз в том случае, если продуценты намного превосходят первичных консументов по своим размерам (примером может служить экосистема леса, где продуцентами являются деревья, а первичными консументами — насекомые).
2) Пирамида биомассы (на схеме — 2). С ее помощью можно наглядно показать соотношения биомасс на каждом из трофических уровней. Она может быть прямой, если размер и срок жизни продуцентов достигает относительно больших величин (наземные и мелководные экосистемы), и обращенной, когда продуценты невелики по размеру и имеют короткий жизненный цикл (открытые и глубокие водоемы).
3) Пирамида энергии (на схеме — 3). Отражает величину потока энергии и продуктивность на каждом из трофических уровней. В отличии от пирамид численности и биомассы, пирамида энергии не может быть обращенной, так как переход энергии пищи на вышестоящие трофические уровни происходит с большими энергопотерями. Следовательно, суммарная энергия каждого предыдущего трофического уровня не может быть выше энергии последующего. Вышеприведеное рассуждение основано на использовании второго закона термодинамики, поэтому пирамида энергии в экосистеме служит его наглядной иллюстрацией.
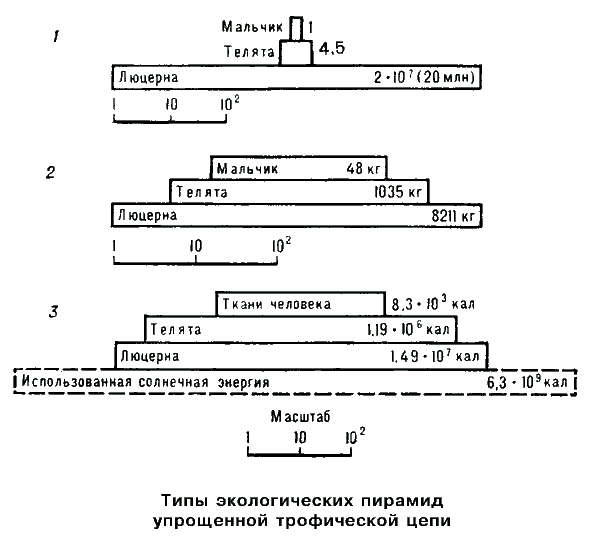
Из всех названных выше трофических характеристик экосистемы только пирамида энергии дает наиболее полное представление об организации биологических сообществ. В пирамиде численности сильно преувеличена роль мелких организмов, а в пирамиде биомассы завышено значение крупных. В таком случае, данные критерии непригодны для сравнении функциональной роли популяции, сильно различающихся по значению отношения интенсивности метаболизма к размеру особей. По этой причине, именно поток энергии служит наиболее подходящим критерием для сравнения отдельных компонентов экосистемы между собой, а также для сравнения двух экосистем друг с другом.
Знание основных законов превращения энергии в экосистеме способствуют лучшему пониманию процессов функционрования экосистемы. Это особенно важно в связи с тем, что вмешательство человека в ее естественую "работу" может привести экологическую систему к гибели. В связи с этим, он должен уметь заранее предсказывать результаты своей деятельности, и представление об энергетических потоках в экосистеме сможет обеспечить большую точность этих предсказаний.