Методическое издание
Александр Петрович Осипов
Вера Михайловна Аксенова
ФИЗИОЛОГИЯ СИСТЕМЫВЫДЕЛЕНИЯ
Методическое пособие для студентов
зооинженерного и ветеринарного факультетов
Подписано в печать. Формат 60х84/16. Бум ВХИ.
Печать на ризографе. Усл.-печ.л., Уч.-изд.л..
Тираж 100 экз. Заказ.
ИПЦ «Прокростъ»
Пермской государственной сельскохозяйственной академии
имени академика Д.Н. Прянишникова
614090, г. Пермь, ул. Коммунистическая, 23 тел. 210-35-34
Ó ФГОУ ВПО «Пермская ГСХА», 2007
| ОГЛАВЛЕНИЕ | Стр. |
| Введение ……………………………………………………………... | |
| 1. ФУНКЦИОНАЛЬНАЯ ХАРАКТЕРИСТИКА ПОЧЕК ………... | |
| 1.1. ВЫДЕЛИТЕЛЬНАЯ ФУНКЦИЯ ПОЧЕК …………………... | |
| 1.1.1. Структурная организация нефрона ……………………... | |
| 1.1.2. Механизмы образования мочи ………………………….. | |
| 1.1.2.1. Клубочковая фильтрация ………………………… | |
| 1.1.2.2. Канальцевая реабсорбция ………………………… | |
| 1.1.2.3. Канальцевая секреция …………………………….. | |
| 1.1.2.4. Синтез выделяемых веществ ……………………... | |
| 1.1.3. Регуляция мочеобразования ……………………………... | |
| 1.1.4. Состав и свойства конечной мочи ………………………. | |
| 1.2. СЕКРЕТОРНАЯ ФУНКЦИЯ ПОЧЕК ……………………….. | |
| 1.3. ГОМЕОСТАТИЧЕСКАЯ ФУНКЦИЯ ПОЧЕК ……………... | |
| 2. МЕХАНИЗМЫВЫВЕДЕНИЯ МОЧИ …………………………. | |
| 3. СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ХАРАКТЕРИСТИКА КОЖИ И ЕЕ ПРОИЗВОДНЫХ …………………………………….. | |
| 3.1. Кожа ……………………………………………………………. | |
| 3.2. Железы ………………………………………………………… | |
| 3.3. Волосы …………………………………………………………. | |
| 3.4. Перья …………………………………………………………… | |
| 3.5. Прочие образования кожи ……………………………………. | |
| 4. Контрольные вопросы для самопроверки знаний ……………… | |
| 5. Вопросы для подготовки к лабораторному занятию …………... | |
| 6. Экзаменационные вопросы ……………………………………… | |
| 7. Список использованных терминов и сокращений ……………... | |
| 8. Список рекомендуемой литературы ……………………………. | |
| 9. Приложение. Эволюция выделительной системы ……………... | |
ВВЕДЕНИЕ
В процессе жизнедеятельности в органах и тканях образуются продукты, которые не могут далее использоваться в обменных процессах. Накопление таких веществ в организме может нарушить гомеостаз. Поэтому одним из механизмов поддержания нормальной жизнедеятельности человека и животных является удаление во внешнюю среду конечных продуктов обмена, чужеродных и находящихся в избытке веществ.
Органы, выводящие вещества из организма объединяют в систему органов выделения. К ним относятся: почки, кожа, слизистая оболочка желудочно-кишечного тракта (ЖКТ), печень, легкие, потовые и сальные железы, пищеварительные железы, носовые и ректальные солевые железы (у птиц и рыб), а также жабры (у рыб).
Через легкие при выдохе выводятся углекислый газ, пары воды и летучие вещества. Пищеварительные железы и слизистая оболочка ЖКТ выделяют тяжелые металлы, ряд лекарственных веществ и чужеродных органических соединений. Печень с желчью удаляет из крови гормоны, продукты обмена гемоглобина, азотистого метаболизма и многие другие вещества. Поджелудочная железа наряду с солями тяжелых металлов выделяет пурины и лекарственные вещества. Через кожу выделяются вода, соли, некоторые органические вещества (например, мочевина, мочевая кислота и молочная кислота).
Однако, важнейшим органом выделения являются почки.
Поэтому понимание и знание функций органов выделения и в первую очередь почек необходимо каждому зооинженеру и ветеринару.
ФУНКЦИОНАЛЬНАЯ ХАРАКТЕРИСТИКА ПОЧЕК
Почки участвуют в выполнении многих функций. Они обеспечивают поддержание постоянства внутренней среды организма, являются эндокринным органом, синтезируют необходимые организму вещества, но безусловно важнейшей является выделительная функция, которая обеспечивает выведение из организма воды и водорастворимых веществ.
1.1. ВЫДЕЛИТЕЛЬНАЯ ФУНКЦИЯ ПОЧЕК
Почка выполняет выделительную функцию благодаря уникальной способности к мочеобразованию. Поскольку большинство веществ, дальнейшее накопление которых может причинить вред организму, являются водорастворимыми, именно с мочой они удаляются из крови. У беспозвоночных функции почек выполняют протонефридии и метанефридии, а у простейших - сократительные вакуоли.
Образование мочи происходит в нефронах, которые являются структурно-функциональными единицами почек.
1.1.1. Структурная организация нефрона
В каждой почке млекопитающих находится около 1 млн нефронов (рис. 1).

| Рис. 1. Схема строения мальпигиева тельца: 1 - приносящая артериола; 2 - выносящая артериола; 3 - капсула Шумлянского - Боумена, 4 - капилляры сосудистого клубочка; 5 - просвет клубочка. Стрелками обозначено направление движения жидкостей (крови в кровеносных сосудах и первичной мочи из клубочка в проксимальный извитый каналец) |
Начальный отдел нефрона (мальпигиево тельце) состоит из клубочка окруженного двумя слоями капсулы Шумлянского-Боумена. Клубочек состоит из капилляров, на которые разветвляется приносящая артериола. При выходе из клубочка капилляры образуют одну выносящую артериолу.
Проксимальный извитый каналец выходит из капсулы Шумлянского - Боумена (рис. 2) и после нескольких изгибов превращается в петлю Генле. Она спускается в вещество почки (нисходящий каналец), а затем поворачивает на 180° и снова приближается к клубочку (восходящий каналец).

| Рис. 2. Схема нефрона: 1 - капсула Шумлянского-Боумена; 2 - проксимальный извитый каналец; 3 - собирательная трубочка, 4 - петля Генле; 5 - дистальный извитый каналец; 6 - Мальпигиев клубочек; 7 - выносящая артериола; 8 - приносящая артериола. Стрелками обозначено направление движения жидкостей (крови в кровеносных сосудах и мочи в канальцах) |
Те нефроны, у которых петли Генле короткие и лишь незначительно проникают в мозговое вещество называются корковыми, а нефроны с глубоко проникающими в мозговой слой длинными петлями Генле - юкстамедуллярными.
В корковом веществе почки петля Генле переходит в извитый дистальный каналец. Удаленный от петли Генле конец дистального извитого канальца соединяется с собирательной трубкой, которая начинается в корковом веществе, а в мозговом слое объединяется с собирательными трубками других нефронов в выводной проток, из которого моча последовательно поступает в почечную лоханку, мочеточники и мочевой пузырь.
Почечная артерия, в почке разветвляется до приносящих артериол, которые снабжают кровью капилляры клубочка (первая капиллярная сеть). Затем капилляры образуют выносящую артериолу, по которой кровь выходит из клубочка и снова распадается на перитубулярные капилляры (вторая капиллярная сеть), окружающие канальцы всех нефронов и петли Генле корковых нефронов. От второй капиллярной сети в мозговое вещество почки параллельно всем собирательным трубкам и петлям Генле юкстамедуллярных нефронов отходят прямые сосуды к почечной вене. Таким образом, особенностью кровообращения в почках является наличие двух капиллярных сетей и прямых сосудов.
Вся кровь в организме животного, проходит через почки за 4-5 мин. Причем, интенсивность почечного кровотока достаточно стабильна. Изменения почечной гемодинамики обычно наступают лишь при интенсивной мышечной нагрузке, под действием адреналина, а также при значительном падении системного артериального давления.
1.1.2. Механизмы образования мочи
В нефроне непрерывно осуществляются четыре необходимых для образования конечной мочи процесса. Ими являются: фильтрация, реабсорбция, секреция и синтез.
Клубочковая фильтрация
Первым этапом образования мочи является фильтрация плазмы крови в мальпигиевых клубочках нефрона. Она приводит к образованию большого объёма первичной мочи,содержащей как ядовитые, так и нужные данному животному преимущественно низкомолекулярные вещества.
Фильтрация не требует затрат АТФ и происходит благодаря диффузии компонентов плазмы крови через поры в стенках капилляров сосудистого клубочка. Размер пор позволяет проходить только таким низкомолекулярным соединениям как глюкоза, аминокислоты, витамины, некоторые гормоны, мочевина, мочевая кислота, креатинин, электролиты и вода. Клетки крови и почти все белки плазмы из-за своих больших размеров остаются в кровеносных сосудах. Благодаря этому в просвете клубочка образуется близкий по составу плазме крови ультрафильтрат.
Силой определяющей скорость клубочковой фильтрации, является фильтрационное давление (ФД). Оно отражает соотношение гидростатического и онкотического градиентов в просвете клубочка и его капиллярах. Гидростатическое давление в капиллярах клубочков (ГД) способствует фильтрации, а градиент онкотического давления (Ронк) и гидростатическое давление жидкости (Ртк) в капсуле противодействуют ей.
Меньший, по сравнению приносящим, диаметр выносящего сосуда затрудняет отток крови из капилляров клубочка и повышает в них ГД до 70-90 мм рт. ст. (в других капиллярах ГД не превышает 40 мм рт. ст.). Ртк в паренхиме почек близко к 10 мм рт. ст. Поэтому градиент ГД и Ртк направлен в сторону просвета клубочков и равен 60-80 мм рт. ст. В оттекающей от клубочков крови Ронк повышено примерно до 35 мм рт. ст., так как в плазме (из-за потери части воды) увеличена концентрация белков, а первичная моча их практически не содержит. Следовательно, Ронк давление в ней близко к нулю, а градиент этого давления направлен в сторону кровеносных капилляров клубочка и близок 35 мм рт. ст.
ФД рассчитывается по формуле ФД = ГД-(Ронк+Ртк) и в норме составляет не менее 30 мм рт. ст. При ФД близком к нулю образование мочи невозможно. Следовательно, при ГД в капиллярах клубочка, равном ГД в капиллярах других органов, первичная моча образоваться не может.
Таким образом, клубочковая фильтрация обеспечивает образование первичной мочи, в которой содержатся практически все растворенные в плазме крови низкомолекулярные вещества. Из каждых 10 л крови, проходящей через капилляры клубочков, образуется около 1 л первичной мочи и только 0,01 л выделяемой организмом вторичной мочи. Следовательно, дальнейшие процессы в нефроне должны обеспечивать возврат (реабсорбцию) в кровь практически 99% объема ультрафильтрата.
Канальцевая реабсорбция
Первичная моча из клубочков поступает в извитые канальцы, петлю Генле и собирательные трубочки. Здесь осуществляется реабсорбция (обратное всасывание) необходимых организму веществ через стенки канальцев. Они выстланы клетками эпителия, в мембранах которого имеются структуры обеспечивающие активный (с затратой энергии АТФ) и пассивный (без затрат энергии) транспорт. Одно и то же реабсорбируемое вещество через одну мембрану клетки канальцев может проходить пассивно, а через другую - активно. Причем, активный транспорт одного вещества может способствовать пассивному перемещению другого. Например, Na+ (рис. 3) из просвета канальцев пассивно входит в клетки по концентрационному (в клетке его мало) и электрохимическому (внутренняя поверхность мембраны клеток электроотрицательна) градиентам. Выведение Na+ из клеток в межклеточные пространства происходит с помощью натриевых насосов (для работы насосов нужна энергия АТФ ). Вслед за Na+ пассивно всасываются анионы бикарбоната, а хлориды абсорбируются пассивно в области контактов клеток проксимальных канальцев. Вместе с ними по осмотическому градиенту реабсорбируется вода.
Наибольшая часть веществ (более 80%) из первичной мочи реабсорбируется в проксимальном извитом канальце. В том числе здесь полностью возвращаются в кровь глюкоза, белки, аминокислоты, витамины и некоторые гормоны, а также около 2/3 воды, Na+, К+, двухвалентных катионов, хлоридов, бикарбонатов, фосфатов и до 50% мочевины.
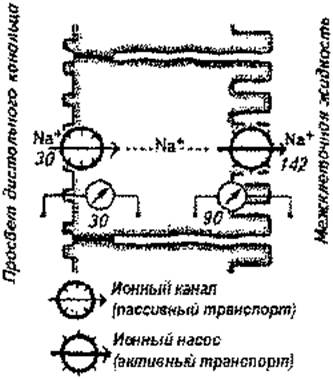
| Рис. 3.Механизмы реабсорбции ионов натрия. |
Проксимальный извитой каналец является самой длинной и широкой частью нефрона. Его стенки состоят из эпителиальных клеток с множеством микроворсинок на внутренней поверхности канальца. Большая площадь внутренней поверхности, обилие митохондрий и близость кровеносных капилляров позволяют проксимальным канальцам интенсивно реабсорбировать вещества из первичной мочи.
Белки и полипептиды, поступившие в просвет почечного канальца, пиноцитируются эпителием проксимальных канальцев и расщепляются до аминокислот, которые используются этими клетками или диффундируют в перитубулярные капилляры.
В проксимальном отделе нефрона неорганические вещества всасываются в результате сопутствующей активному транспорту Na+, пассивной реабсорбции бикарбоната и воды, а также пассивным транспортом Cl- с пассивной реабсорбцией Na+ и осмотически эквивалентного объема воды. Поэтому к петле Генле подходит жидкость изоосмотичная плазме крови.
Реабсорбция глюкозы и аминокислот обеспечивается общим переносчиком с Na+. Образовавшийся комплекс пассивно (по электрическому и концентрационному градиентам) поступает в клетку, где распадается с освобождением Na+ и глюкозы (или аминокислоты). Затем из клетки в межклеточное пространство Na+ переносится с помощью натриевого насоса, а глюкоза и аминокислоты диффундируют по градиентам концентраций.
Избыток глюкозы в крови (и следовательно в ультрафильтрате) вызывает нехватку ее переносчиков и излишек глюкозы выделяется с мочой. Таким образом, для глюкозы канальцевая реабсорбция ограничена «почечным порогом выведения». При его достижении глюкоза и другие «пороговые вещества» выводятся с вторичной мочой.
У млекопитающих и птиц имеется противоточно-множительная канальцево-сосудистая система (состоит из петли Генле, собирательных трубочек и расположенных параллельно канальцам кровеносных капилляров) снижающая затраты воды на выведение водорастворимых веществ.
Данная система создает в мозговом веществе почек нарастание осмотического давления по направлению от границы коркового вещества к почечному сосочку. Это способствует интенсивному удалению воды из канальцев и приводит к накоплению в собирательной трубке гиперосмотичной (по отношению к плазме крови) мочи.
Работа противоточно-множительной канальцево-сосудистой системы обусловлена разной проницаемостью и особенностями расположения ее отделов.
Верхний сегмент нисходящей части петли Генле непроницаем для солей, мочевины и воды. По нему фильтрат переходит из проксимального извитого канальца в нижний (длинный и тонкий) сегмент нисходящей части петли Генле, который пропускает лишь воду. Поэтому вода (по осмотическому градиенту) выходит из нисходящей части петли Генле в межклеточную среду, а затем вместе с растворенными в ней ионами - в прямые кровеносные сосуды. При этом в восходящую часть ее поступает концентрированная жидкость.
Нижний, тонкий сегмент восходящей части петли Генле хорошо проницаем для хлористого натрия и мочевины, но не проницаем для воды. Здесь по концентрационным градиентам хлористый натрий выходит из канальцев, а мочевина поступает в них. В толстом сегменте восходящей части петли эпителий активно переносит ионы натрия и хлора из фильтрата. Следовательно, в восходящем отделе петли Генле реабсорбируются тольк ионы. Это приводит к росту осмотического давления в межклеточной жидкости окружающей оба отдела (находятся близко друг к другу) данной петли, а в дистальные канальцы поступает гипотоничный фильтрат.
Концентрированию мочи в петле Генле способствуют узкие нисходящие и широкие восходящие прямые кровеносные сосуды. Они отделены от канальцев тонким слоем межклеточного матрикса. Поэтому растворенные вещества сначала выходят из петли в интерстиций мозгового вещества почки. В нисходящем прямом сосуде из плазмы крови по градиенту осмотического давления в тканевую жидкость выходит вода, а по концентрационным градиентам в кровь диффундируют хлористый натрий и мочевина. Это увеличивает осмотическое давление плазмы крови в сосуде по мере его приближения к колену петли Генле. В восходящем сосуде вода поступает в кровь, а ионы выходят из нее. Поэтому осмотическое давление вытекающей из почек крови нормализуется, а в тканевой жидкости почек повышается, что способствует реабсорбции воды из канальцев.
Таким образом, в петле Генле реабсорбируются: вода, Na+ и Cl-. Образующаяся при этом у птиц и млекопитающих моча значительно уменьшается в объеме и становится гиперосмотичной.
Корковые нефроны поддерживают постоянное содержание воды в кровеносном русле только при достаточном поступлении ее в организм. При дефиците воды возрастает роль юкстамедуллярных нефронов, способных снижать её потери за счет значительного увеличения концентрации выводимых веществ в моче.
Дистальные извитые канальцы приближаются к мальпигиеву тельцу в корковом веществе почки. Именно здесь регулируется водно-солевой баланс и рН крови. Эндотелий дистальных канальцев активно реабсорбирует К+, Са++, фосфаты и Na+, а Cl- перемещается пассивно вслед за Na+. Одновременно с этим, в обмен на ионы натрия дистальные канальцы удаляют из межклеточных пространств в мочу ионы водорода, что способствует поддержанию КЩР в организме.
Канальцевая секреция
Механизмы канальцевой секреции обеспечивают выведение из организма недостаточно фильтрующихся или не поступающих в первичную мочу веществ. К ним относятся некоторые ионы (например, К+ и Н+), органические кислоты, эндогенные основания, чужеродные (например, антибиотики, рентгеноконтрастные вещества и красители) и другие вещества. Органические соединения и ионы водорода секретируются преимущественно в проксимальных, ионы калия - в дистальных, а аммиак - в проксимальном и дистальном извитых канальцах.
Клетки эпителия канальцев захватывают из межклеточной жидкости подлежащие удалению вещества и секретируют их в его просвет. Этому способствуют находящиеся в эндотелии и на его поверхностях ферменты. Например, эффективное удаление из крови ионов водорода обеспечивается активностью карбоангидразы на апикальной мембране клеток канальцев. Данный фермент ускоряет образование угольной кислоты из двуокиси углерода и воды. После этого угольная кислота быстро диссоциирует, а появившиеся в клетке ионы водорода секретируются в просвет канальца.
Интенсивность секреции возрастает по мере увеличения необходимости удаления тех или иных компонентов, а также зависит от концентрации в моче и плазме других веществ. Например, секреция К+ усиливается высокой концентрацией карбонатов и сульфатов в канальцах.