И движения везикул
Везикулы образуется путем почкования мембран. Различают три способа образования везикул. В зависимости от вида почкующейся мембраны и направления транспорта везикулы:
1 - образуются от мембраны одной органеллы (донора) и переме- щаются к мембранам другой органеллы (акцептора) (везикулярный транспорт белков и липидов, синтезируемых в ЭПС);
2 - образуются от плазмолеммы и перемещаются к месту транспор- та (эндоцитоз);
3 - образуются от мембран органелл и перемещаются к плазмалем-ме и выводятся из клетки (экзоцитоз).
По достижению места транспорта происходит слияние мембраны пузырька с мембранной полости акцептора. При этом происходит высвобождение содержимого пузырька в полость акцептора. В случае слияния с плазмолеммой (экзоцитоз), высвобождение содержимого пузырька происходит вне клетки. Эти процессы сопровождаются перестройкой органелл или поверхности клетки с сохранением или разрушением межклеточных контактов.
Движение веществ от плазмолеммы в клетку и от аппарата Гольджи к плазмолемме обеспечивают транспортные пузырьки, имеющие диа- метр 500А и окруженные решетчатой оболочкой, состоящей из белка клатрина (рис.30А,Б) Клатрин состоит из трех крупных и трех менее крупных полипептидов. Все субъединицы объединяются в структуру, называемую трискелион (от греч. skelоs - нога), имеющую вид трехножки (рис.30,В) с «бедрами» и «голенями» длиной около 25 нм. Трискелионы соединяются латерально вдоль своих «конечностей», образуя шестигранную ячейку, но не плоской, а куполообразной формы; ячейки собираются вместе, образуя решетчатую структуру.

Рис.30.Окаймленный клатриновый пузырек: А - схематическое изображение пузырка; Б - общий вид на электронограмме; В - сборка клатриновой сети из субъединиц в виде «трехножек»: 1- крупные полипептиды, 2 – менее крупные полипептиды (Из: Фаллер, Шилдс, 2003; Де Дюв, 1987)
Мембрана в месте расположения клатриновой решетки начинает выпячиваться (рис.31А,Б), формируя окаймленную ямку. Затем края ямки смыкаются с образованием окаймленного пузырька. Пузырек образуется в местах высокой концентрации белков и липидов внутри ЭПС и содержит высококонцентрированный раствор белков. Таким же путем формируются транспортные окаймленные пузырьки на плазмолемме в процессе эндоцитоза (рис.31 А).
Такие транспортные пузырьки называется окаймленными и переносят мембранные, секреторные и лизосомальные белки от ЭПС к аппарату Гольджи и дальше в целевые акцепторные органеллы. Окаймленные пузырьки транспортируют белки и липиды от плазмалеммы к внутренним мембранам: транспортные белки, факторы роста, инсулин, цитокины в другие части клетки.

Рис. 31. Схема (А) и моде-ль (Б) образования клатриново- го пузырька: 1-3 - этапы образо-вания клатринового пузырька; 4 – клатриновый пузырек (Из: Де Дюв, 1987; Фаллер, Шилдс, 2004)
Образование пузырьков не приводит к изменению донорской, акцепторной мембраны или плазмолеммы, так как осуществляется механизм возвращения мембран (см. «Биология клетки:ЭПС и комплекс Гольджи.»), при котором часть белков и липидов мембран пузырьков возвращается к месту образования пузырьков. В случае возврата везикул от аппарата Гольджи к ЭПС этот процесс называется ретроградным транспортом. Если грузовые вещества пузырька продолжают перемещается к месту транспорта, то такой транспорт называют антероградным. Раздельный транспорт грузовых молекул и компонентов мембраны обеспечивается двумя классами белков со специфичической каймой. Эти белковые комплексы называется коатомеры или СОРS (рис.32). Различают коатомеры СОР-I и СОР-II. СОР-I состоит из 8 субъединиц и обеспечивают ретроградный транспорт, СОР-II состоит из 5 субъединиц и обеспечивает антероградный транспорт и перенос пузырьков от ЭПС к аппарата Гольджи.
В процессе везикулярного транспорта главную роль играют молекулы белков, так называемых молекулярных переключателей. Они являются рецепторами мембраны пузырьков и активизируют молекулы-мишени мембран органелл-акцепторов, после активации которых происходит присоединение СОРS к мембране.
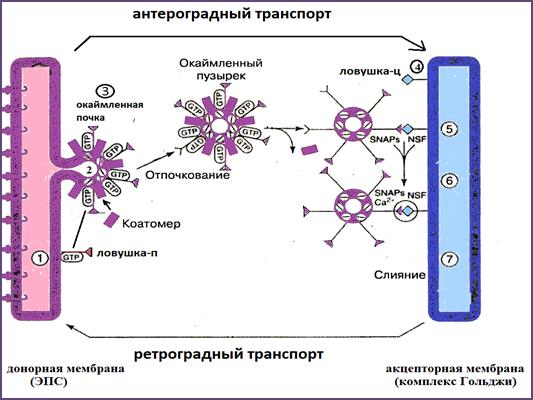
Рис. 32. Антероградный и ретроградный транспорт, осуществляемый транс - портными окаймленными пузырьками: 1 – Ловушка-п в составе мембраны донора; 2,3 – образование почки пузырька, сопровождающееся присоединением коатомеров I; 4 – транспорт окаймленного пузырька к акцепторной мембране; 5 – Ловушка-ц; 5,6 – участие белков SNAPs и NSF в присоединении окаймленного пузырька к акцепторной мембране; 7 – слияние мембран пузырька и акцептора(Из:Фаллер, 2003)
Для обеспечения правильного направления транспорта пузырьков, каждый пузырек снабжен специальной рецепторной меткой, называемой ЛОВУШКА-п (для пузырька); целевая мембрана снабжено рецептором, называемым ЛОВУШКА -ц (целевая мембрана). Рецептор ЛОВУШКИ-п связывается с рецептором ЛОВУШКИ-ц, обеспечивая, правильную доставку пузырьков.
Слияние мембраны пузырька с целевой мембранной происходит при участии специального белка слияния (фузионный белок); одним из них является Rаb-белок, который обеспечивает направление движения транспортных пузырьков. Для каждой внутриклеточной мембраны существует своя разновидность Rаb-белка.
Везикулярный транспорт обеспечивает следующие клеточные процессы: проведение электроимпульса по нервным клеткам, секрецию гормонов, производство компонентов крови, клеточный иммунитет, развитие эмбриона, формирование органов, внутриклеточный обмен веществ. По направлению движения везикулярного транспорта и по харак-теру переносимых веществ различают следующие процессы: эндоцитоз и экзоцитоз.
Эндоцитоз
Эндоцитоз – везикулярный перенос жидкостей, макромолекул и небольших твердых частиц вместе с частью плазмалеммы из внешней для клетки среды в клетку. В зависимости от направления транспорта и характера транспортируемых веществ различают 3 типа механизмов эндоцитоза: пиноцитоз, фагоцитоз и рецепторно-опосредованный эндоцитоз или клатрин-зависимый эндоцитоз.
Независимо от типа механизма, любой эндоцитоз проходит по одному плану (рис.33):
1 - образуется впячивание в месте контакта плазмалеммы с поглощаемым веществом с образованием ямки;
2 - ямка углубляется, её края слипаются и смыкаются; образуется пузырек – везикула, в полости которого находится поглощенное вещество;
3 - пузырек отщепляется и проникает внутрь цитоплазмы клетки.
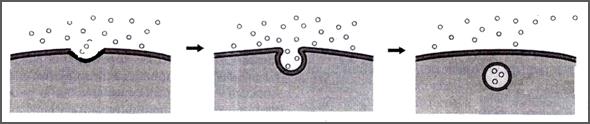 Рис.33. Механизм эндоцитоза. Объяснение в тексте (Из: Фаллер, 2003)
Рис.33. Механизм эндоцитоза. Объяснение в тексте (Из: Фаллер, 2003)
Пиноцитоз (от греч. «пино » - пью и «цитос » - клетка)- это процесс переноса воды и растворенных в ней внеклеточных макромолекулярных соединений через плазмалемму и поглощение их клеткой. В месте захвата поглощаемые вещества неспецифически воздействуют на рецепторы мембраны. Это воздействие передается на подмембранный слой микрофиламентов и далее на цитоскелет, который вызывает впячивание плазмалеммы в виде тонкого канальца или ямки (рис.34).

Рис.34. Процесс образования пиноцитозных пузырьков: А – образование пиноцитозного канала (1), отпочковывание от канала пиноцитозных пузырьков (2), пиноцитозные пузырьки, отпочковывающиеся от ямки; Б – образование ямки (1) и пузырька(2) (Из: Интернет-ресурсов с сайта: biologiamedica.blogspot.com GIF 800×600)
В них и попадает жидкость с растворенными в ней веществами. От канальца затем отпочковываются пузырьки; они отщепляются от мембраны и проникают внутрь клетки. Если образуется ямка, то она углубляется, края её слипаются, а затем смыкаются и отщепляется пузырек (рис. 35Б-2)
Несмотря на малые размеры пиноцитозных пузырьков, их многочисленность позволяет доставлять в клетку большое количество веществ. Пиноцитоз может быть очень интенсивным процессом; в некоторых клетках вся плазматическая мембрана поглощается и восстанавливается за один час. Пиноцитозные пузырьки, содержащие макромолекулы, в клетке сливаются и образуют ранние эндосомы. которые, сливаясь с первичными лизосомами, превращаются в поздние эндосомы или вторичные лизосомы. В них и происходят процессы расщепления (рис.35).
Пиноцитозные пузырьки могут оставаться в клетке, но могут мигрировать в противоположную от места образования сторону клетки и там, встраиваясь в плазмалемму, выбрасывать свое содержимое из клетки. Этот процесс называется трансцитоз (рис. 36). Он обеспечивает транспорт веществ, поступивших в клетку извне к различным местам внутри клетки, в другие клетки, а также в клетки других слоев ткани. Особенно активно он протекает в клетках кровеносных и лимфатических сосудов. Таким образом, транспортируются иммуноглобулины матери через молоко в организм ребенка.

Рис.35. Процесс обра-зования вторичных лизо-сом при участии пиноци-тозных пузырьков ((Из: Интернет-ресурсов с сай-та health. mail.ru )

Рис. 36. Трансцитоз (Из: Ин- тернет-ресурсов с сайта health. mail.ru)
Фагоцитоз (от греч. «фагос » - пожирать и «цито с » - клетка) – это захват клетками относительно крупных нерастворенных в воде (0,5 мкм) частиц и высокомолекулярных соединений с помощью актин-зависимого механизма. Клетки, обеспечивающие подобный захват называются фагоцитами.
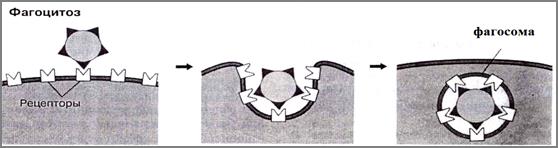
Рис.37. Схема фагоцитоза. Объяснение в тексте (Из: Фаллер, 2003)
Фагоцитоз включает в себя восемь этапов (рис. 38):

Рис. 37. Схема фагоцитоза (Из: Ярилин A.A., 20)
1 – хемотаксис, который обеспечивает приближение фагоцита к микробной клетке. Хемотаксис - это движение клеток в сторону высокой концентрации химических факторов: хемотоксинов или хемоаттрак- тантов, которые выделяются микроорганизмами или активизируются клеткой в месте осуществления фагоцитоза, например, в очаге воспаления (цитокины, лейкотриен В4, гистамин) (рис.38,1);
2 - прилипание фагоцита к объекту поглощения (рис.38,2). Мембрана фагоцита несет специфичные рецепторы на своей поверх- ности и своеобразные химические замочки, с помощью которых микроорганизм пристегивается к фагоциту.
3 - активация мембраны фагоцита под действием С-протеинкиназы. Осуществляется подготовка к погружению поглощаемого объекта в клетку, важным моментом которой выступает процесс полимеризации актина подмембранного слоя клеточной оболочки. Для запуска этого процесса молекулы поглощенной частицы взаимодействуют с поверхностными рецепторами фагоцита (например, Fс-рецептор). Рецептор генерирует сигнал, который активизирует близлежащие молекулы гиалоплазмы (например,srс-киназа), те, в свою очередь, фосфорилируют цитоплазматическую часть поверхностного Fс-рецептора. Сигналы фосфорилирования полимеризуют G-актин (см. Биология клетки: структура цитоскелета), что приводит к формированию F-актина. F-актин необходим для образования ложноножек, окружающих поглощаемую частицу (рис. 38,3).
4 - погружение частиц в фагоцит. При погружении частицы в цитоплазму, соприкасающаяся с ней часть мембраны впячивается в клетку. Образуются ложноножки, которые обволакивают частицу, таким образом, что вокруг объекта образуется оболочка из фагоцита (рис.38, 4);
5 - образуется везикула, состоящая из крупных частиц, окруженных плазмолеммой, которая называется фагосома (рис.38,5).
6 - фагосома сливается с первичной лизосомой, содержащей ферменты - гидролазы, расщепляющие жиры, белки и углеводы (рис.38,6). В результате образуется фаголизосома или вторичная лизосома. При слиянии мембран фагосомы и лизосомы происходит изменение струк- туры мембраны фагосомы: рецепторы белка (М-6-Ф-рецептор и Fс-рецептор возвращаются назад к плазмалемме, взамен им мембрана фагосомы приобретает другие молекулы (например, Ловушки), которые облегчают слияние мембран фагосомы и лизосомы.
7 - в фаголизосоме происходит разрушение поглощенных частиц под действием гидролаз (рис.38,7).
8 - фагоцитоз завершается выбросом остатков объекта, не приносящих организму вреда (рис.38,8). Объектом фагоцитоза способны выступить вирусы, бактерии, грибки, а также другие частицы, не являющиеся генетически родственными данному организму.
У многоклеточных организмов фагоцитоз обеспечивает:
1 - механизм защиты организма-хозяина от микроорганизмов;
2 - механизм обновления тканей и заживления ран путем поглоще- ния поврежденных и постаревших клеток.
Для микроорганизмов фагоцитоз является защитой от антител и цитотоксичных веществ.
Фагоцитоз млекопитающих осуществляют клетки 3 типов: нейтрофилы, моноциты и макрофаги. Рецепторы этих клеток распознают иммуноглобулины и другие молекулы иммунной системы.
Крупные частицы или внедрившиеся бактерии сначала окружаются иммуноглобулинами (опсонинами) (рис.38). Этот процесс называется опсонизацией. Опсонинами являются вещества, обеспечивающие связывание бактерий и антигенов фагоцитами. К ним относятся антитела (IgA, IgG), лектин, связывающий маннозу М-6-Ф- рецептора, фибронектин и др.
Рис. 38. Процесс опсонизации, предшествующий фагоцитозу (Из: Фаллер,2003)
Рецепторы фагоцита распознают иммуноглобулин и связываются с ним и начинается процесс фагоцитоза (рис.39). Процесс сопровождается выработкой фагоцитом сигнальных молекул (радикалы О2 и N2), ферментов, разрушающих внеклеточный матрикс и другие компоненты, мобилизирующие процесс воспаления.
Кроме этих, основных фагоцитов, фагоцитирующей способностью обладают фибробласты, поглощающие коллаген; клетки эпителия сетчатки, захватывающие палочки и др.

Рис. 39. Схема фагоцитоза, осуществляемый макрофагом с помощью опсонино
а - фиксация опсонинов на поверхности поглошаемой частицы, б – обра- зование прочной связи поглошаемой частицы с макрофагом, в - образование лож- ноножек и погружение поглошаемой частицы внутрь макрофага, г - перемещение частицы внутрь макрофага и ее "переваривание" (Из: Ярилин A.A., 20)
Рецепторно-опосредованный эндоцитоз – это эндоцитоз, при котором поглощаемое с поверхности клетки вещество (лиганд) предварительно связывается со специфическими рецепторами плазмолеммы (рис.40) и транспортируется в клетку в составе окаймленного пузырька. При этом обеспечивается определенные преимущества перед другими видами эндоцитоза:
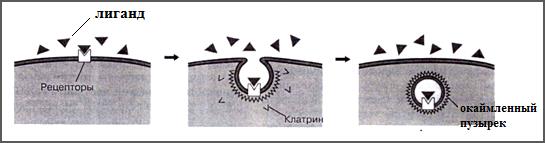
Рис. 40. Схема рецепторно-опосредованного эндоцитоза (Из: Фаллер, 2003)
1 - поглощаются только определенные молекулы и обеспечивается их избирательное связывание;
2 - происходит более быстрое поглощение лиганда в комплексе с рецепторами клетки;
3 - возможность удаления определенного лиганда из внеклеточной среды.
Рецепторно-опосредованный эндоцитоз осуществляется с помощью клатриновых везикулярных пузырьков, называемых также окаймленными (см.рис.30) и начинается со связывания лиганда с рецеп- тором мембраны, который находится в определенным месте поверх- ности, называемом окаймленная ямка (рис.41 из фаллер стр. 109).
Поверхность окаймленной ямки со стороны цитоплазмы покрыта клатрином. За связыванием следует образование окаймленного пузырька. Он входит в клетку и сливается там с первичной лизосомой с образованием вторичной лизосомы. Далее, в зависимости от характера лиганда и рецептора рецепторно-опосредованный эндоцитоз может пойти по следующим путям:
1 - рецептор возвращается к плазмалемме, лиганд разрушается во вторичной лизосоме;
2 - лиганд-рецепторный комплекс возвращается к плазмалемме;
3 - лиганд-рецепторный комплекс разрушается в лизосоме; такой путь осуществляется в процессе обеспечения клеточного ответа, в случае необходимости удаления рецептора с поверхности клетки;
4 - лиганд-рецепторный комплекс транспортируется через клетку и доставляется на её другую сторону.
Рецепторно – опосредованный эндоцитоз обеспечивает поступление в клетку:
1-транспортных белков: ЛНП (липопротеины низкой плотности), транспортирующий холестером и трансферрин, траснпортирующий железо;
2- факторы роста (соматотропный гормон роста, фактор роста тромбоцитов), инсулин, цитокины - вещества, обусловливающие клеточный ответ (подробнее см.ниже: «Передачи сигналов в клетку»);
3-вирусы и токсины;
4- пептидные гормоны, иммунные комплексы;
5-циторецепторы;
6-желточные включения в овоците и ряд других веществ.
Нарушения рецепторно-опосредованного эндоцитоза приводят к ряду серьезных заболеваний. Одно из них семейная гиперхолестеролемия. Это заболевние обусловливается повышением уровня ЛНП в плазме крови в связи с нарушением связывания холестерола с рецептором ЛНП. Это нарушение вызывается мутацией гена рецептора ЛНП; при этом рецептор ЛНП не может связаться с холестеролом и он не попадает в клетку. Такой ген называется рецепторно-негативным. Другая мутация этого гена вызывает недостаточность поступления холестерола, третья мутация этого гена нормально связывает рецептор и холестерол, но нарушается процесс перемещения комплекса лиганд-рецептор к окаймленной ямке.
Рецепторно-опосредованный эндоцитоз является одним из вариан- тов пино- и фагоцитоза в иммунных процессах.
Экзоцитоз
Экзоцитоз - это транспорт крупных соединений и частиц из клетки. В зависимости от направления транспорта веществ различают следую- щие типы экзоцитоза:
1 - секреция: выделение клеткой растворимых продуктов секреции или обмена веществ;
2 - экскреция: выделение твердых частиц,
3 - рекреция: поглощение твердых частиц из вне клетки, их транс- порт через клетку и выделение наружу.
Выделяемые из клетки путем экзоцитоза вещества могут оставаться на ее оболочке в виде рецепоров, могут входить в состав межклеточного вещества или выполнять роль сигнальных молекул (нейротрансмиттеры, гормоны). Секреторные и рекреторные пузырьки перемещаются в результате сокращения цитоскелета к плазмолемме, сливаются с ней и секрет оказывается за пределами клетки.

Рис.41. Типы экзоцитоза. Объяснение в тексте (Из: Мушкамбарова,2003)
Экзоцитоз в клетке осуществляется в результате следующих этапов:
1 - транспортировка везикулы от места синтеза и формирования (аппарат Гольджи) до плазмалеммы. Транспорт обеспечивают моторные белки динеин и кинезин вдоль актиновых филаментов либо микротрубочек цитоскелета.
2 - стыковка везикулы с плазмалеммой клетки путем прочного соединения белковых компонентов обеих мембран, при участии особых белковых комплексов.
3 - объединение липидного бислоя везикулы с бислоем плазма - леммы и слияние мембраны везикулы с плазмалеммой клетки. После окончания слияния мембран происходит высвобождение или выброс содержимого секретируемой везикулы во внеклеточное пространство. Секреторная функция - секреция веществ на «экспорт» осуществляется в процессе 5 стадий:
1 - поглощение извне клетки веществ, которые служат для синтеза веществ «на экспорт». Эта фаза обеспечивается транспортными системами клетки: пиноцитозными пузырьками, фагосомами и ионными каналами;
2 - синтез веществ «на экспорт» в гранулярной ЭПС и комплексе Гольджи;.
3 - накопление синтезированного секрета в клетке в виде секре -торных пузырьков и гранул (см.ниже).
4 - выделение секрета при помощи мерокринового (без разруше - ния клеток), апокринового (с разрушением части клетки) или голокринового (с разрушением всей клетки) механизмов секреции.
5. Восстановление первоначального состояния клетки.
Путем секреции выделяются из клетки высококомплексные соединения (например, гормоны гипофиза) и низкомолекулярные соединение (ионы Н+, активные амины, медиаторы и т.д.).
Различают конститутивную и регулируемую секрецию.
Кальций-независимая к онститутивная секреция – это постоянный процесс выделения веществ без внешних для клетки сигналов и наличия кальция. Встречается практически во всех эукариотических клетках. Во многих клетках синтезируется пептидные гормоны, пищеварительные ферменты и другие белки, которые постоянно перемещаются в составе секреторных пузырьков от комплекса Гольджи (белки на «экспорт») к плазмолемме и выделяется наружу. Кроме того, такой тип секреции используется в процессе построения внеклеточного матрикса и доставки белков на внешнюю клеточную мембрану. Секреторные везикулы доставляются к поверхности клетки и сливаются с наружной мембраной.
Кальций-зависимая регулируемая секреция – это процесс выделения веществ с помощью внешних сигналов, гормональных или нервных, например, при выделения нейромедиаторов. При этом типе секреции секреторные вещества накапливаются в секреторных пузырьках в виде крупных гранул. Пузырьки не сливаются с плазматической мембраной и не высвобождают свое содержание до тех пор, пока клетка не получит внешний сигнал, который вызывает приток ионов кальция через плазмалемму и быстро повышает концентрацию ионов кальция в цитоплазме клетки. В пресинаптических мембранах такой процесс осуществляет кальций-зависимый белковый комплекс SNARE.
Если, в секреторных пузырьках накапливаются вещества, например белки, в количествах превышающих их концентрацию в ЭПС примерно в 200 раз, то образуются плотные гранулы, которые называются секреторными гранулами с плотным ядром (рис.42 из фаллер, стр.93).
Экзоцитоз обеспечивает выполнение трех основных задач:
1 - доставка к плазмалемме клетки липидов, необходимых для её роста;
2 - выведение из клетки различных соединений: токсичных продук- тов обмена веществ, белков и липидов на «экспорт» или сигнальных молекул (гормонов, нейромедиаторов и пр.);
3 – доставка к плазмалемме рецепторов или транспортных белков.
Механизмы экскреции и рекреции веществ подобны механизмам секреции, но отличаются содержимым везикул.
Везикулярный транспорт происходят в клетках непрерывно. При этом объем мембран, совершающих оборот, значителен: например, в течение 1 часа макрофаг поглощает в виде везикул двойной объем своей поверхности своей.
Нарушение везикулярного транспорта является причиной возникновения ряда заболеваний: муковисцидоз, наследственная эмфи -зема легких, болезнь I-клеток и др.