Тема: «Типы скрещиваний»
Вопросы.
1. Классификация типов скрещиваний в селекции древесных растений.
2. Простые скрещивания.
3. Сложные скрещивания.
Ключевые слова:
скрещивания, простые скрещивания, сложные скрещивания, топкросс, поликросс, парные скрещивания, диаллельные скрещивания, прямые скрещивания, обратные скрещивания, реципрокные скрещивания, гибриды, простые гибриды, сложные гибриды, ступенчатые скрещивания, возвратные скрещивания, насыщающие скрещивания, конвергентные скрещивания, межгибридные скрещивания, множественные скрещивания, однократные скрещивания, многократные скрещивания.
Распределение бюджета рабочего времени
Форма проведения занятий – аудиторные занятия, лекция.
Бюджет времени – 2 часа.
Количество двухчасовых занятий – 1.
Распределение бюджета времени:
- 1 час: рассмотрение существующих теоретических представлений о типах скрещиваний, применяемых в лесной селекции;
- 1 час: рассмотрение проблем и перспектив применения простых и сложных скрещиваний я в лесной селекции.
Литература.
1. Гужов Ю.Л., Фукс А., Валичек П. Селекция и семеноводство культурных растений. – М.: Агропромиздат,1991. – 463 с. – (Учебники и учеб. Пособия для студентов высш. Учеб. Заведений).
2. Рокицкий П.Ф. Введение в статистическую генетику. Минск: Вышейшая школа, 1978. – 448 с.
3. Бессчетнов В.П., Кентбаев Е.Ж., Бессчетнова Н.Н, Кентбаева Б.А. Лесная селекция / В.П. Бессчетнов, Е.Ж. Кентбаев, Н.Н Бессчетнова, Б.А Кентбаева. - Учебник для студентов лесохозяйственных специальностей. Гриф Республиканского учебно-методического совета Республики Казахстан. – Алматы: Нур-Принт, 2015. – 358 с.
4. Бессчетнова Н.Н. Сосна обыкновенная (Pinus sylvestris L.). Селекционный потенциал плюсовых деревьев / Н.Н. Бессчетнова. – Saarbrücken: Lap Lambert Academic Publishing GmbH & co. KG. ISBN 978-3-8443-5608-3, 2011 – 402 c.
5. Бессчетнова Н.Н., В.П. Бессчетнов. Сосна обыкновенная (Pinus sylvestris L.). Морфометрия и физиология хвои плюсовых деревьев / Н.Н. Бессчетнова, В.П. Бессчетнов. Монография. – Нижний Новгород: Нижегородская государственная сельскохозяйственная академия, 2014. – 368 с.
6. Бессчетнова Н.Н. Сосна обыкновенная (Pinus sylvestris L.). Репродуктивный потенциал плюсовых деревьев / Н.Н. Бессчетнова. Монография. – Нижний Новгород: Нижегородская государственная сельскохозяйственная академия, 2015. – 586 с.
7. Бессчетнова Н.Н. Сосна обыкновенная (Pinus sylvestris L.). Эффективность отбора плюсовых деревьев / Н.Н. Бессчетнова. Монография. – Нижний Новгород: Нижегородская государственная сельскохозяйственная академия, 2016. – 382 с.
8. Бессчетнов В.П. Полиморфизм казахстанских популяций облепихи крушиновидной (Hippophae rhamnoides L.) по хозяйственным и адаптивным признакам. Дис. док. биол. наук. Алматы, 1994. – 543 с.
9. Бессчетнов В.П. Полиморфизм казахстанских популяций облепихи крушиновидной (Hippophae rhamnoides L.) по хозяйственным и адаптивным признакам. Дис. док. биол. наук. Алматы, 1994. – 543 с.
10. Бессчетнов В.П.
11. Чмыр А.Ф., Бессчетнов В.П. Экология и культура облепихи Hippophae rhamnoides L. – СПб, СПбНИИЛХ, 1998. – 278 с.
12. Джонатан В. Райт. Введение в лесную генетику. М.: Лесная промышленность, 1978. – 470 с.
13. Котов М.М. Генетика и селекция. Часть 1.: Учебник для вузов. – Йошкар-Ола: МарГТУ, 1997. – 280 с.
14. Котов М.М. Генетика и селекция. Часть 2.: Учебник для вузов. – Йошкар-Ола: МарГТУ, 1997. – 108 с.
15. Любавская А.Я. Лесная селекция и генетика.: Учебник для вузов. – М.: Лесная промышленность, 1982. – 288 с.
Асинхронно-синхронная связь с преподавателем:
1. Бессчетнов Владимир Петрович: E-mail: lesfak@bk.ru
Мобильный телефон: 8-906-355-81-97.
2. Бессчетнова Наталья Николаевна: E-mail: besschetnova1966@mail.ru
Мобильный телефон: 8-960-160-54-32.
Классификация типов скрещиваний в селекции древесных растений
Все типы скрещиваний деревьев и кустарников можно подразделить на две основные группы: простые и сложные (Любавская, 1982). При этом подразумевают то, что в результате простых скрещиваний образуются простые гибриды, а в результате сложных скрещиваний – сложные гибриды.
Простые гибриды – это гибриды, в генотипе которых объединена наследственность только двух исходных родительских форм.
Сложные гибриды – это гибриды, в генотипе которых объединена наследственность более двух исходных родительских форм.
Схематически структура и взаимоотношения этих категорий скрещиваний может быть представлена следующим образом (рис. 1).
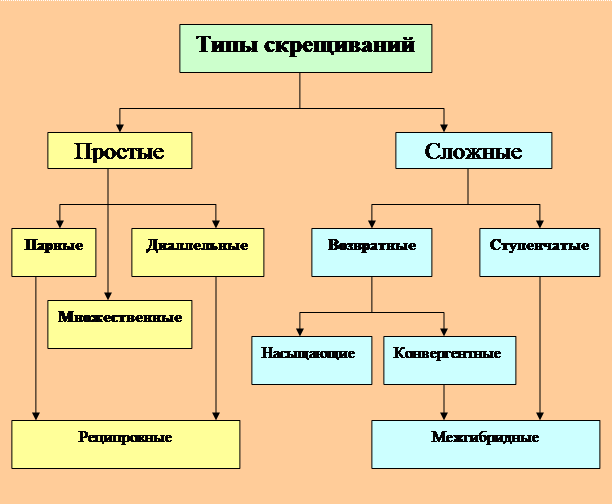
Рисунок – 1. Схема типов скрещиваний
(реконструкция по А.Я. Любавской, 1982 с коррекцией В.П. Б.)
В литературе (Гужов, Фукс, Валичек, 1991, стр.171) приводятся и другие классификации типов скрещиваний. Согласно таким подходам все скрещивания делят на две группы: однократные и многократные. Здесь также следует понимать, что однократные скрещивания подразумевают объединение в гибриде как в конечном результате синтетической селекции наследственности только двух родительских форм, например, только двух плюсовых деревьев. Многократные скрещивания в своем итоге – полученном гибриде – предусматривают объединение наследственности нескольких (более двух) исходных родительских форм – носителей признаков, которые должны быть у гибрида, в соответствии с целью его получения. Схематически это можно изобразить следующим образом (рис. 2; по Гужову, Фуксу, Валичеку, 1991, стр.171).

Рисунок – 2. Схема типов скрещиваний (реконструкция по Гужову, Фуксу, Валичеку, 1991, стр.171 с коррекцией Б.В.П.)
И хотя данная классификация типов скрещиваний, на мой взгляд, не вполне совершенна, в ней есть одно существенное обстоятельство – то, что множественные скрещивания совершенно справедливо отнесены к простым скрещиваниям. В других учебниках (Любавская, 1982) множественные скрещивания необоснованно отнесены к сложным. Тем не менее, такой подход к организации селекции методом гибридизации при практическом проведении скрещиваний широко известен.
Простые скрещивания.
Наиболее широкое распространение в лесной селекции получили простые скрещивания.
Простые скрещивания – это скрещивания между двумя родительскими формами, производимые однократно.
Такое определение вы найдете во всех, или, по крайней мере, в большинстве учебников по селекции растений или лесной селекции. Вместе с тем оно не отражает основное содержание простых скрещиваний.
Простые скрещивания – это скрещивания, в результате которых в генотипе гибридов объединяется наследственность только двух исходных родительских форм.
Понятно, что такой результат возможен только в случае, когда в процессе получения гибридов скрещиваются только две родительские формы, а само скрещивание осуществляется однократно.
Собственно говоря, все типы скрещиваний: и диаллельные, и сложные, – являются лишь различными сочетаниями, схемами объединения и комбинаций простых скрещиваний. Простые парные скрещивания – конструкционная основа всех остальных типов скрещиваний. Это определено биологией процесса оплодотворения – процесса слияния гамет – возможно только однократное слияние в зиготу только одной пары гамет – одного спермия и одной яйцеклетки.
Простые скрещивания с той или иной степенью условности подразделяют на парные и диаллельные. Разница между ними состоит в схеме организации гибридного процесса.
Простое парное скрещивание – это собственно однократное скрещивание межу двумя родительскими формами.
Относительно видов деревьев и кустарников оно может быть реализовано как на однодомных так и на двудомных растениях. Схематически простое парное скрещивание может быть отражено следующим образом.
♀А × ♂В = F(AB)
Формообразовательный процесс в гибридных популяциях от простых скрещиваний идет на основе перераспределения наследственного материала, привнесенного в равном количестве одной парой родителей.
Диаллельное скрещивание – это система простых парных скрещиваний, при реализации которой формы, линии, сорта, плюсовые или элитные деревья скрещиваются между собой во всех возможных комбинациях (полные диаллельные скрещивания) или только в части комбинаций (неполные диаллельные скрещивания) (Гуляев, Мальченко, 1975, стр. 166).
И простые парные скрещивания, и диаллельные скрещивания, могут быть прямыми и обратными, а при объединении той и другой их схемы – реципрокными.
Прямое простое парное скрещивание.
♀А × ♂В = F(AB)
Обратное простое парное скрещивание.
♀В × ♂А = F(BA)
Реципрокное простое парное скрещивание.
♀А × ♂В = F(AB)
♀В × ♂А = F(BA)
Реципрокные скрещивания – это скрещивания между двумя родительскими формами, когда каждая из них в одном случае берется в качестве материнского, а в другом – в качестве отцовского родителя.
Понятно, что в абсолютном смысле это относится к однодомным видам – большинству видов деревьев и кустарников – каждая особь которых способна выступать и в качестве материнского организма (на котором образуются женские гаметы, а позже образуются и разовьются зиготы, завязи и семена), и в качестве отцовского организма (который будет источником мужских гамет – источником пыльцы).
В подавляющем большинстве случаев при реализации реципрокных схем скрещиваний признаки и свойства гибрида не зависят от направления скрещивания. Объясняется это тем, что как в прямом, так и в обратном скрещивании ядерный материал, контролирующий развитие всех признаков организма (при ядерной наследственности), привносится гаметами в зиготу в равных количествах.
В случаях же, когда развитие каких-либо признаков контролируется цитоплазмой, между реципрокными гибридами будут наблюдаться существенные различия: признак будет передаваться только в том скрещивании, где материнской формой берется форма с цитоплазматической наследственностью, определяющей наличие данного признака.
Кроме того, от выбора материнского родителя часто зависит процент завязываемости гибридных семян при внутривидовых скрещиваниях и в особенности при отдаленных (например, межвидовых) скрещивания.
Учитывая то, что эндосперм семени при отсутствии двойного оплодотворения формируется из гаплоидных клеток материнского организма,…при возвратном скрещивании качество семян может быть отличающимся от генетической характеристик семян, полученных от прямого скрещивания.…
Реципрокные скрещивания применяют в следующих основных случаях (Царев и др., 2005, стр. 73).
1. Когда наследование какого-либо важного хозяйственно-биологического признака (например, зимостойкости, роста и др.) связано с цитоплазмой; гибриды наследуют это свойство сильнее в том случае, если носитель ценного признака берется в качестве материнского растения.
2. Когда завязываемость семян зависит от того, в качестве материнской или отцовской берется та или иная форма.
диаллельные скрещивания применяются преимущественно для оценки небольшого числа линий, форм, сортов, плюсовых или элитных деревьев по специфической комбинационной способности, после их предварительной оценки по общей комбинационной способности в скрещиваниях по системе топкросс (Гуляев, Мольченко, 1975, стр. 166). Для получения оценок СКС необходимы контролируемые скрещивания. При работе с лесными древесными и кустарниковыми видами эти селекционные мероприятия долговременны и весьма дороги. Многовариантные схемы особенно затратны. В этой связи при организации селекционных мероприятий, направленных на проведение контролируемых скрещиваний между плюсовыми или элитными деревьями, предварительно составляют схему (план) скрещиваний. При их разработке предусматривают достижение наибольшей рациональности таких схем, их разумную минимизацию и эффективность, возможность достижения нескольких целей. При этом учитывают то, что существует ряд основных схем диаллельных скрещиваний:
- полная схема диаллельных скрещиваний;
- модифицированная схема диаллельных скрещиваний;
- частичные схемы диаллельных скрещиваний;
- факториальная схема скрещиваний;
- простые скрещивания.
Полная схема диаллельных скрещиваний (рис. 2.1) считается одной из лучших в смысле широты получаемых результатов, но и наиболее сложной в практическом выполнении.
| Матери ♀ | Отцы ♂ | |||||||||
| × | × | × | × | × | × | × | × | × | × | |
| × | × | × | × | × | × | × | × | × | × | |
| × | × | × | × | × | × | × | × | × | × | |
| × | × | × | × | × | × | × | × | × | × | |
| × | × | × | × | × | × | × | × | × | × | |
| × | × | × | × | × | × | × | × | × | × | |
| × | × | × | × | × | × | × | × | × | × | |
| × | × | × | × | × | × | × | × | × | × | |
| × | × | × | × | × | × | × | × | × | × | |
| × | × | × | × | × | × | × | × | × | × |
Рис. 2.1. Полная схема диаллельных скрещиваний, включающая варианты самоопыления
В ней предусматриваются все возможные варианты скрещиваний, и как следствие получается почти полная информация о генетических характеристиках изучаемых объектов (например, клонов плюсовых деревьев). Совокупность всех скрещиваний (и прямых, и обратных) в данной схеме представляет собой реципрокные скрещивания. Она способна дать информацию об ОКС и СКС и их дисперсиях. Результат, получаемый при реализации данной схемы (семенное потомство плюсовых деревьев известного происхождения), представляет собой оптимальный материал для отбора из числа испытываемых плюсовых деревьев лучших индивидуумов (по оценке ОКС) или пар клонов (по оценке СКС), подходящих для создания биклоновых или моноклоновых лесосеменных плантаций (ЛСП). Нетрудно заметить, что схема будет очень сложной в практической реализации, прежде всего по экономическим соображениям. Так в случае её реализации для 10 клонов, как на приведенном рисунке, потребуется провести 100 контролируемых скрещиваний, а для 20 – 400. В общем случае для такой схемы скрещиваний число их возможных комбинаций будет n × n = n2. Учитывая то, что каждый вариант скрещивания проводится в нескольких (3 – 10) повторностях, нетрудно понять, что число собственно контролируемых опылений будет еще многократно больше. Кроме того, каждая повторность каждого варианта скрещивания будет представлена некоторым набором сеянцев (10 – 50), что также приведет к существенному увеличению учетных растений при последующем изучении полученных потомств. Поэтому на практике в лесной селекции используют другие схемы.
Вариантом полной схемы диаллельных скрещиваний при работе с перекрестно опыляющимися видами, к которым относятся практически все виды деревьев и кустарников, является схема, в которой исключены все варианты самоопыления (рис. 2.2).
| Матери ♀ | Отцы ♂ | |||||||||
| × | × | × | × | × | × | × | × | × | ||
| × | × | × | × | × | × | × | × | × | ||
| × | × | × | × | × | × | × | × | × | ||
| × | × | × | × | × | × | × | × | × | ||
| × | × | × | × | × | × | × | × | × | ||
| × | × | × | × | × | × | × | × | × | ||
| × | × | × | × | × | × | × | × | × | ||
| × | × | × | × | × | × | × | × | × | ||
| × | × | × | × | × | × | × | × | × | ||
| × | × | × | × | × | × | × | × | × |
Рис. 2.2. Полная схема диаллельных скрещиваний, исключающая варианты самоопыления
Число возможных вариантов ней составит n × (n – 1).
Модифицированная схема диаллельных скрещиваний (рис. 2.3) представляет собой ограниченную схему диаллельных скрещиваний (её определенный фрагмент).
| Матери ♀ | Отцы ♂ | |||||||||
| × | ||||||||||
| × | × | |||||||||
| × | × | × | ||||||||
| × | × | × | × | |||||||
| × | × | × | × | × | ||||||
| × | × | × | × | × | × | |||||
| × | × | × | × | × | × | × | ||||
| × | × | × | × | × | × | × | × | |||
| × | × | × | × | × | × | × | × | × |
Рис. 2.3. Модифицированная схема диаллельных скрещиваний (только прямые односторонние скрещивания, исключая варианты самоопыления)
При её реализации из полной схемы исключают все обратные скрещивания и самоопыления, в силу чего схема перестает быть реципрокной. Само по себе это значительно упрощает схему, делает её легче выполнимой, менее затратной и приводит к удешевлению работ. При этом данная схема дает примерно такую же по объему и содержанию информацию, как и полная схема диаллельных скрещиваний, но ограничения, допущенные в ней, не гарантируют такой же точности в опытах при определении параметров. Число возможных вариантов простых парных скрещиваний в ней будет равно n × (n – 1) / 2.
Частичная схема диаллельных скрещиваний (рис. 2.4) может значительно отличаться от полной и модифицированной схем диаллельных скрещиваний, прежде всего тем, что часть вариантов скрещиваний вообще исключается из схемы их реализации. Сокращение возможных вариантов скрещивания может быть настолько значительным, что отдельный клон не скрещивается со всеми другими.
| Матери ♀ | Отцы ♂ | |||||||||
| × | × | |||||||||
| × | × | |||||||||
| × | × | |||||||||
| × | × | |||||||||
| × | × | |||||||||
| × | × | |||||||||
| × | × | |||||||||
| × | × | |||||||||
| × | × | |||||||||
| × | × |
Рис. 2.4. Частичная схема диаллельных скрещиваний
Даная схема менее эффективна при получении оценок ОКС и СКС, чем полная и модифицированная схемы диаллельных скрещиваний. Отсутствие оценок, которые нельзя получить в частичных схемах, значительно усложняет расчеты. Однако это в той или иной степени компенсируется тем, что большое количество потомства может быть испытано при сравнительно низких затратах. В представленном варианте схемы число возможных комбинаций (прямых и обратных) будет 4n.
Близкой к рассмотренным выше схемам простых диаллельных скрещиваний является факториальная схема скрещиваний.
факториальная схема скрещиваний (рис. 2.5) – это схема скрещиваний, в которой каждая из всех материнских форм скрещивается с одним и тем же набором отцовских форм (клонов плюсовых деревьев) и наоборот, которые в случае однодомных деревьев и кустарников не выступают в качестве материнских особей.
В случае, когда в указанную схему скрещиваний вовлечены двудомные растения (тополя, ивы, облепиха или другие), базовое определение факториальной схемы скрещиваний воспринимается однозначно и не требует пояснений, дополнений или уточнений. В вариантах использования однодомных видов (сосна, дуб, ель, лиственница и т.д.), каждая особь которых в схемах скрещиваний может выступать как женской, так и мужской родительской формой, определение нуждается в уточнении, приведенном выше.
| Матери ♀ | Отцы ♂ | ||||
| × | × | × | × | × | |
| × | × | × | × | × | |
| × | × | × | × | × | |
| × | × | × | × | × | |
| × | × | × | × | × |
Рис. 2.5. Факториальная схема скрещиваний
Понятно, что в этой схеме число материнских особей может быть и больше, и меньше числа отцовских форм (или равно ему).
Данная схема может рассматриваться как разновидность полной диаллельной схемы, включающей в себя все комбинации одной группы матерей и другой группы отцов (рис. 2.6).
| Матери ♀ | Отцы ♂ | |||||||||
| × | × | × | × | × | ||||||
| × | × | × | × | × | ||||||
| × | × | × | × | × | ||||||
| × | × | × | × | × | ||||||
| × | × | × | × | × |
Рис. 5. Факториальная схема скрещиваний в системе диаллельных скрещиваний.
Эта схема была широко распространена в селекционных программа США под названием “Северо-Каролинская II”.
Преимущества данной схемы состоят в её сравнительной простоте и относительной дешевизне, а также в том, что анализ результатов легче.
Вместе с тем имеются и заметные недостатки. Поскольку в схеме, как правило, используется ограниченное количество отцовских особей (клонов), и в связи с тем, что одни и те же клоны (особи) не являются одновременно и материнскими, и отцовскими, трудно сравнить (дать надежную оценку) ОКС каждого родителя, задействованного схеме. Также трудно отобрать потомство для следующего поколения лесосеменных плантаций.
ПРИМЕЧАНИЯ
В методику – к обоснованию целесообразности применения скрещиваний наиболее отдаленных (по своим характеристикам) форм. В учебнике Царева и др. (2005, стр.79) со ссылкой на С.С. Пятницкого (1961, стр. …) отмечается, что «… при скрещивании материалов из географически отдаленных»
[]
[Пятницкий С.С. Практикум по лесной селекции. М.: Изд-во сельскохозяйственной литературы, журналов и плакатов, 1961. – 271 с.]
Географическая отдаленность и происхождение исходных форм из разных в экологическом смысле условий произрастания подразумевает то, что эти формы эволюционно (в смысле приспособления к разным экологическим условиям и изоляции) различаются, а, следовательно, различны и их генотипы. В соответствии с этим, поиск наиболее различающихся по своим признакам и свойством объектов (клонов плюсовых деревьев) является важной и вполне конкретной селекционной задачей, решение которой обеспечит больший успех селекции вида в целом.
Географическая отдаленность и происхождение исходных форм из разных в экологическом смысле условий произрастания – это лишь пути формирования (причины, сформировавшие) наибольших по своим проявлениям генетических различий между сравниваемыми и скрещиваемыми особями (клонами ПД). В тоже время факт состоит в том, что объекты имеют различия в генотипе, выраженные в той или иной степени, и в том, что, чем выше степень генетических различий, тем больший положительный эффект (по мнению, С.С. Пятницкого) ожидается от их скрещивания.
При множественных скрещиваниях (поликроссах) опыление какого либо (одного) материнского растения осуществляется смесью пыльцы нескольких отцовских форм. Схематически это можно изобразить так (В.П.).
(А+А+А+А+….+А) × (В+С+D+Е + … +Z) =
= (AB)+(AC)+(AD)+(АЕ)+ … +(AZ)
Схемы из учебников (Любавская, 1982; Гужов, Фукс, Валичек, 1991) содержат логическую неточность (указана толь одна буква А), приводящую к неправильному восприятию смыслового содержания данной схемы скрещиваний. Кроме того, в них не акцентируется внимание на то, что результатом реализации схемы будет комплекс – множество – гибридов, различающихся генотипами отцов и относящихся друг к другу как полусибсы.
Приведенная мною схема отличается от тех, что представлены практически во всех учебниках селекции. В ней я посчитал правильным представить материнскую форму так, как именно она участвует в данной схеме скрещиваний – множеством женских гамет. Это соответствует и смысловому содержанию данной схемы, и её устоявшемуся названию.
Здесь буквами «А+А+А+А+ … +А » обозначена материнская форма (клон, выступающий в качестве материнской особи с множеством генеративных органов или тем более комплекс рамет одного ортета), буквами «В+С+D+Е+ … + …+Z)» обозначены отцовские формы (клоны других плюсовых деревьев, выступающие в качестве опылителей).
Понятно, что результатом такого скрещивания будет множество простых парных гибридов, поскольку само множественное скрещивание выступает как некоторый комплекс простых парных скрещиваний какого-либо клона, представленного множеством рамет, или даже одного дерева с множеством цветков или макростробилов. В генотипе каждого отдельного гибрида будет объединен генотип материнской формы и генотип одного (только одного!) из мужских растений (клонов плюсовых деревьев – опылителей).
Его уместнее было бы представить схемой, в результате реализации которой получается комплекс – множество – простых гибридов.
A × B = AB
A × C = AC
A × D = AD
A × E = АЕ
……………
……………
A × Z = А Z
Если внимательно рассмотреть данную схему, то становится очевидным, что она адекватна фрагменту схемы свободного опыления растений на изолированной многоклоновой ЛСП, когда в процессе опыления каждый отдельный клон будет опылен пыльцой всех остальных клонов, введенных в состав ЛСП. Собственно фрагментом схемы выступают все варианты комбинаций одного клона, например: плюсового дерева с неким индексом «А» (ПД-А), - со всеми остальными клонами плюсовых деревьев. При этом, конечно же, недопустимо присутствие посторонней пыльцы деревьев, не входящих в состав ЛСП. Приведенная схема может быть рекомендована как для однодомных видов (сосна, ель, дуб, ясень и др.), так и для двудомных (тополя, ивы, облепиха и др.). В случае однодомных видов при полной свободе опыления мы получим полную диаллельную схему скрещиваний, в которой семенное потомство каждой отдельной материнской формы (семенное потомство всех рамет одного клона, выступающего как материнская форма) будет по сути дела потомством, полученным по схеме множественных скрещиваний, не исключающей самоопыления (см. схему на рис. 2.6).
| Матери ♀ | Отцы ♂ | |||||||||
| × | × | × | × | × | × | × | × | × | × | |
Рис. 2.6. Множественные скрещивания особи «1», содержащие самоопыление
Или при отсутствии самоопыления…
| Матери ♀ | Отцы ♂ | |||||||||
| × | × | × | × | × | × | × | × | × | ||
Рис. 2.7. Множественные скрещивания особи «1», не содержащие самоопыления
При включении в данный процесс (данную схему скрещиваний) клонов всех пл