Липиды не образуют макромолекул, аналогичных белкам, полисахаридам или нуклеиновым кислотам – полимерам, образованным ковалентно связанными повторяющимися структурными единицами. Однако благодаря своей амфифильности липиды способны образовывать высокоорганизованные структуры, соизмеримые с макромолекулами. Вместе с белками такие структуры образуют бислойные биологические мембраны, которые окружают все клетки и служат своеобразными перегородками между ними и внутри клеток.
Все биологические мембраны, включая плазматическую мембрану и внутренние мебраны эукариотических клеток, имеют общие структурные особенности: они представляют собой ансамбли липидных и белковых молекул, удерживаемые вместе с помощью нековалентных взаимодействий. Благодаря этим взаимодействиям поддерживается структурная целостность мембран.
Плазматическая мембрана, окружающая клетку, определяет ее величину и обеспечивает сохранение существующих различий между клеточным содержимым и окружающей средой. Мембрана – это не просто барьер, а активно работающее сложное устройство:
1. Служит высокоизбирательным фильтром и, кроме того, отвечает за активный транспорт; с ее помощью регулируется поступление внутрь клетки питательных веществ и выход наружу продуктов выделения. Благодаря мембране устанавливается разница в концентрации ионов внутри клетки и во внеклеточном пространстве.
2. Как следствие трансмембранного градиента концентраций ионов (в первую очередь калия и натрия) возникает трансмембранный электрический потенциал, служащий источником энергии для осуществления клеткой полезной работы (см. раздел 2.8).
3. С мембранами (в первую очередь митохондриальными) связаны каскады ферментов, что позволяет осуществлять последовательную передачу промежуточных продуктов с одного фермента на другой.
4. Плазматическая мембрана содержит специфические рецепторы для внешних возбудителей или для сигналов других клеток, органов и тканей у многоклеточных (гормоны, маркеры, антигены и т.д.). Взаимодействие с рецептором приводит к изменению свойств мембраны (например, вязкости), активации встроенных в мембрану ферментов и, в конечном счете, вызывает ответ клетки: например, направленное движение (хемотаксис), генерацию химических или электрических сигналов. Это позволяет клетке быстро отвечать на изменения, происходящие в окружающей среде (одноклеточные) или в организме (многоклеточные).
5. На плазматической мембране расположены специфические для клетки антигены, позволяющие организму отличать ее от больных (раковых) или чужеродных клеток.
Благодаря инвагинациям плазматическая мембрана может образовывать единое целое с протяженными каналами, которые «вводят» окружающую среду далеко «внутрь» клетки и участвуют в формировании пространства между внутренней и внешней мембранами ядра. Эта система каналов находится в постоянной взаимосвязи с внеклеточным окружением.
Плазматические мембраны представляют собой двуслойные липидные структуры толщиной от 6 до 10 нм. Второй основной компонент – белки, составляющие от 20 (эритроциты) до 80% (митохондрии) массы мембраны. Главные мембранные липиды – фосфоглицериды, гликолипиды и холестерин. Однако в различных мембранах липидный состав существенно отличается:
| Эритроциты | Гепатоциты | E.coli | |
| Лецитин (фосфатидилхолин) | 31% | 18% | - |
| Кефалин (фосфатидилэтаноламин) | 15% | 11% | 80% |
| Фосатидилсерин | 7% | 9% | - |
| Сфингомиелин | 8,5% | 14% | - |
| Холестерин | 24% | 30% | - |
Мембранные белки разделяют на интегральные (внутренние) и периферические (внешние). Как интегральные, так и поверхностные белки имеют полярные и неполярные области (домены). Неполярная область погружена в липидную мембрану, полярные области выдвинуты в водное окружение. Периферические белки лишь частично погружены в липидный бислой, значительная часть поверхности их глобулы образована полярными участками. Они легко экстрагируются водными растворами, не содержащими поверхностно-активных веществ. Интегральные белки практически полностью расположены в неполярной области мембраны, часто пронизывая ее насквозь. Внутренние белки нерастворимы в водных растворах, их экстракция возможна только при добавлении поверхностно-активных веществ. Интегральные белки обладают функциональной активностью только в случае погружения в мембрану – только в этих условиях приобретается необходимая конформация (третичная структура). Большинство мембранных белков – ферменты и транспортные системы.
При обычной температуре липидный бислой представляет собой двухмерную жидкость: липиды свободно перемещаются в плоскости мембраны, но «перескок» из одного слоя в другой очень затруднен. Таким образом, мембранные белки могут свободно перемещаться в плоскости мембраны. Возбуждение клетки приводит к перераспределению белков и сборке их в ансамбли. Однако подвижность некоторых белков ограничена или отсутствует за счет специальных структурных образований – микрофиламентов и микротрубочек.
Мембрана асимметрична, что определяется различным липидным и белковым составом наружного и внутреннего слоя и асимметрией встроенных интегральных белков. Кроме того, на внешней поверхности расположено большое количество олигосахаридов, отсутствующих на внутренней поверхности.
Мембранный транспорт
Мембраны легко проницаемы для воды и растворимых в липидах веществ, непроницаема для ионов и полярных неионизированных веществ (например, для сахаров). Следовательно, для переноса последних требуются специальные механизмы.
Способы транспорта через клеточную мембрану:
1. Простая диффузия по градиенту концентрации.
2. Облегченная диффузия по градиенту концентрации.
| В тех случаях, когда простая диффузия не может обеспечить необходимую скорость транспорта. Белки пермеазы и транслоказы избирательно связываются с веществом и переносят его через мембрану. Пример – вход глюкозы в гепатоциты, эритроциты и другие клетки. |

3. Активный транспорт – позволяет переносить вещества против градиента концентрации, для чего необходимо расходование энергии. Энергия может поступать за счет гидролиза АТФ (первично активный транспорт) или за счет одновременного переноса другого вещества по градиенту концентрации (вторично активный транспорт).
| Типичный пример первично активно транспорта – работа Na, К-зависимая АТФазы, о которой уже говорили (раздел 2.8). При каждом цикле работы насоса расходуется одна молекула АТФ, 2 иона калия переносится в клетку и 3 иона натрия – из клетки. То есть за цикл работы из клетки переносится один положительный заряд. В результате внутренняя часть клетки оказывается отрицательно заряженной относительно внешней среды, возникает трансмембранный электрический потенциал около 100 мВ. Учитывая малую толщину мембраны, напряженность электрического поля оказывается очень высокой около 10 000 В/см. На поверхности одного эритроцита расположено 100-200 молекул Na, К-зависимая АТФазы, на поверхности активно функционирующих клеток – до миллиона. |

Са-АТФаза при гидролизе одной молекулы АТФ переносит наружу 2 иона кальция.
В мембранах митохондрий расположен белок, который при добавлении АТФ способен перекачивать протоны (ионы Н+). На самом деле, как увидим в дальнейшем, функция этого белка обратная – синтез АТФ из АДФ и неорганического фосфата при переходе протонов через мембрану по градиенту концентрации.
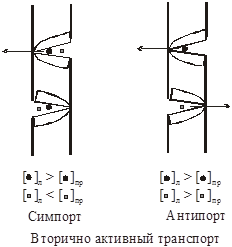
При вторично активном транспорте перенос одного вещества против градиента концентрации оказывается возможен за счет переноса другого по градиенту концентрации. Различают симпорт и антипорт.