Лабораторное занятие № 1.
Оогенез. Строение яйцеклеток и гонад.
Началу индивидуального развития предшествует возникновение половых клеток, т.е. гаметогенез, который можно рассматривать прогенезом при индивидуальном развитии.
Процесс развития женских половых клеток называется овогенезом (оогенезом). В отличие от сперматогенеза он имеет некоторые особенности.
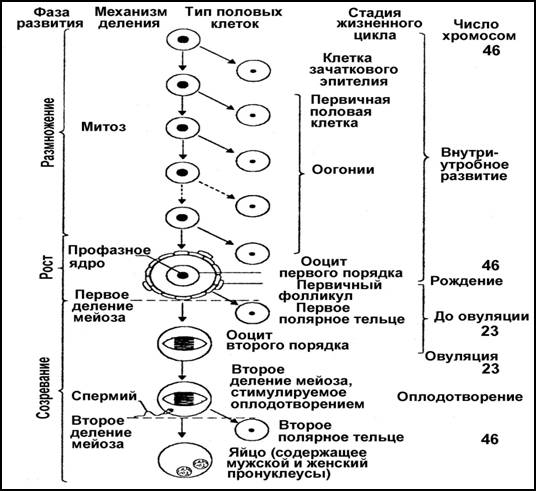
В оогенезе различают 3 периода: размножение, рост и созревание.
1) Недифференцированные женские половые клетки – оогонии – размножаются путем обычного митоза. После деления они становятся ооцитами I порядка и переходят в период роста.
2) Рост ооцитов длится очень долго – недели, месяцы и даже годы. В периоде роста различают два этапа: малый, или медленный рост, когда ассимилируются новые вещества и ими обогащается преимущественно цитоплазма, и большой, или быстрый рост (вителлогенез), когда в клетке накапливается питательный желток. Глубокие изменения в периоде роста претерпевает и ядро, оно сильно набухает, содержимое его как бы расплывается. Размеры клеток колоссально возрастают (например, икринки окуня увеличиваются почти в миллион раз).
Затем ооцит I порядка вступает в период 3) созревания, или мейоз. Здесь совершаются редукционное и эквационное деления. Процессы деления в ядре протекают так же, как при мейозе сперматоцитов, но судьба цитоплазмы совершенно иная. При редукционном делении одно ядро увлекает с собой бóльшую часть цитоплазмы, а на долю другого остается лишь незначительная ее часть. Поэтому образуется только одна полноценная клетка – ооцит II порядка, и вторая крошечная – направительное, или редукционное, тельце, которое может делиться на два редукционных тельца.
При втором, эквационном делении несимметричное распределение цитоплазмы повторяется и опять образуется одна крупная клетка – оотида и третье полярное тельце. Оотида по составу ядра и функционально является вполне зрелой половой клеткой.
Период формирования, в отличие от сперматогенеза, в овогенезе отсутствует!!! Таким образом, в овогенезе из одной овогонии возникает только одна зрелая яйцеклетка. Полярные тельца остаются недоразвитыми и вскоре погибают и фагоцитируются другими клетками. Зрелые женские гаметы называют яйцеклетками или яйцами, а отложенные в воду – икрой.
Кроме этого (!!!), в фазе созревания оогенеза существует 2 блока мейоза:
1. От момента рождения до начала полового созревания – блок профазы 1-го деления мейоза. Снимается благодаря регулирующему действию гормонов гипоталамо-гипофизарно-половой оси.
2. Блок метафазы 2-го деления мейоза – снимается в момент оплодотворения.
Гормональная регуляция оогенеза
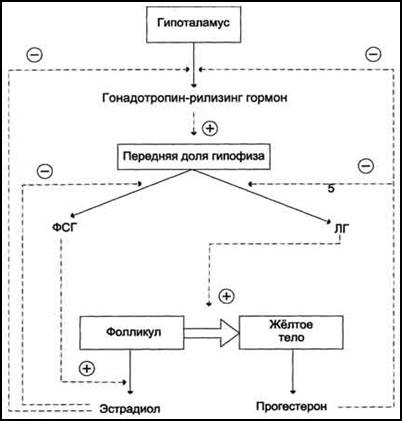
В детском возрасте незрелые яичники вырабатывают небольшое количество гормонов, поэтому концентрация эстрогенов в крови низкая. В пубертатный период чувствительность гипоталамо-гипофизарной системы к действию ЛГ и ФСГ снижается.
У половозрелых женщин гонадолиберин, или гонадотропин-рилизинг-гормон (ГнРГ), секретируется ритмически (пульсирующе) в виде пиков, продолжающихся несколько минут и повторяющихся через 60-90 мин, в ответ на сигналы разнообразных нейромедиаторов. Это, в свою очередь, приводит к циклической секреции ФСГ и ЛГ из передней доли гипофиза. Соотношение секретируемых гормонов (ЛГ:ФСГ) меняется в течение менструального цикла. Следовательно, их высвобождение зависит также от других факторов (гормонов), кроме ГнРГ.
Секреция ФСГ и ЛГ зависит от состояния центральной нервной системы (психогенные факторы, стресс), которая управляет разнообразными медиаторами, с кровотоком поступающими в область гипоталамуса; например, это может быть норадреналин (НА) и нейропептид Y (НП-Y), а также гормоны яичников, эстрогены (эстрон, эстрадиол, эстриол и т. д.), прогестерон и ингибин.
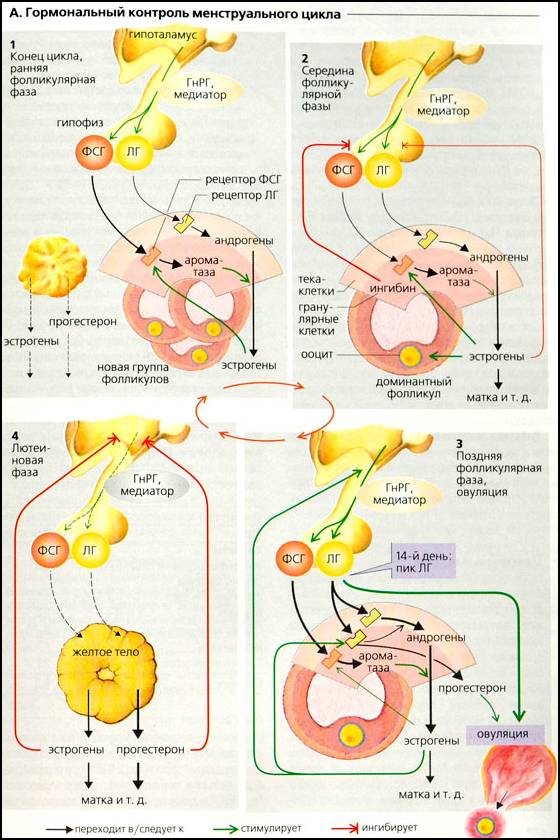
Синтез ФСГ снова увеличивается к концу лютеиновой фазы. В начале фолликулярной фазы ФСГ индуцирует пролиферацию зернистого слоя примерно в 20 фолликулах и стимулирует секрецию ароматазы в их зернистых клетках. Ароматаза катализирует превращение андрогенов – тестостерона и андростендиона – в эстрадиол (E2) и эстрон (Е1). Эстрогены синтезируются в клетках теки и абсорбируются в зернистых клетках. Несмотря на то, что секретируется относительно небольшое количество ЛГ, его достаточно для активации ферментов тека-клеток, которые помогают синтезировать андрогены, необходимые для синтеза эстрогенов. Эстрогены фолликулярного происхождения увеличивают плотность собственных рецепторов ФСГ. Фолликулы с более высокой концентрацией эстрогена, следовательно, являются более чувствительными к ФСГ. Эта петля имеет эффект самоусиления, и конкретный фолликул выбирается в качестве доминантного фолликула примерно на 6-е сутки цикла. В середине фолликулярной фазы эстрогены уменьшают секрецию ФСГ и ЛГ (по системе отрицательной обратной связи и с помощью ингибина), но позже стимулируют продукцию рецепторов ЛГ в зернистых клетках. Эти клетки теперь начинают также синтезировать прогестерон (начало лютеинизации), который абсорбируется тека-клетками и используется в качестве предшественника для дальнейшего увеличения синтеза андрогенов.
Ингибин и эстрогены, секретируемые доминантным фолликулом, ингибируют секрецию ФСГ, таким образом снижая продукцию эстрогенов в других фолликулах. Это ведет к накоплению андрогенов и апоптозу остальных фолликулов.
Увеличивающиеся количества ЛГ и ФСГ высвобождаются в поздней фолликулярной фазе, что вызывает резкое возрастание их концентрации в плазме. Пик ФСГ, наблюдающийся на 13-е сутки цикла, индуцирует первое мейотическое деление ооцита. Эстрогены увеличивают секрецию ЛГ (в основном с помощью гипоталамуса), что приводит к повышенной продукции андрогенов и эстрогенов (положительная обратная связь) и быстрому росту концентрации ЛГ (выброс ЛГ). Пик ЛГ имеет место примерно на 14-е сутки. Фолликул разрывается и высвобождает свою яйцеклетку примерно через 10 часов (овуляция). Овуляции не происходит в том случае, если нет пика ЛГ или если пик слишком мал. При отсутствии овуляции беременность невозможна.
Лютеиновая фаза. ЛГ, ФСГ и эстрогены превращают фолликул ооцита в желтое тело. Оно активно секретирует большие количества прогестерона, начиная лютеиновую фазу. Эстрогены и прогестерон теперь ингибируют секрецию ЛГ и ФСГ как прямо, так и косвенно (например, через ингибирование ГнРГ, см. ранее), что вызывает быстрое падение их концентрации в плазме. Эта отрицательная обратная связь ведет к значительному снижению концентрации в плазме эстрогенов и прогестерона к концу менструального цикла (примерно на 26-е сутки), что вызывает менструальное кровотечение. Секреция ФСГ начинает расти прямо перед началом менструации.
Препарат 5: Яичник кролика
 Изучить корковое вещество яичника, зарисовать стадии развития фолликула и отметить:
Изучить корковое вещество яичника, зарисовать стадии развития фолликула и отметить:
1) целомический эпителий;
2) белочная оболочка;
3) соединительная ткань яичника;
4) примордиальный фолликул (ооцит, окружённый более, чем одним слоем фолликулярных клеток – первичный фолликул);
5) вторичный фолликул
6) третичный фолликул (Граафов пузырёк);
7) тека фолликула;
8) атретический фолликул;
9) фолликулярная жидкость;
10) ооцит;
11) желтое тело;
12) кровеносный сосуд;
13) яйценосный бугорок;
14) лучистый венец;
* * *
Характеристика яйцеклеток:
ü По количеству желтка различают следующие виды яйцеклеток:
1. Полилецитальные — содержат большое количество желтка (членистоногие, рептилии, птицы, рыбы, кроме осетровых).
2. Мезолецитальные — содержат среднее количество желтка (осетровые рыбы, амфибии).
3. Олиголецитальные — содержат мало желтка (моллюски, иглокожие).
4. Алецитальные — не содержат желтка (млекопитающие, некоторые паразитические перепончатокрылые).
ü По распределению желтка в цитоплазме различают следующие виды яйцеклеток:
1. Телолецитальные — желток смещён к вегетативному полюсу яйцеклетки. Противоположный полюс называется анимальным. Сюда относятся некоторые полилецитальные (рыбы, кроме осетровых, рептилии, птицы) и все мезолецитальные яйца (осетровые рыбы, амфибии).
2. Гомо(изо)- лецитальные — желток распределён равномерно. Сюда относятся олиголецитальные ядра (моллюски, иглокожие).
3. Центролецитальные — желток расположен в центре яйцеклетки. Сюда относятся некоторые полилецитальные яйца (членистоногие). Это совершенно особый тип яиц. Вместо анимального и вегетативного полюсов у этих яиц говорят о переднем и заднем полюсах. В центре яйца расположено ядро, а по периферии — ободок свободной от желтка цитоплазмы. Оба этих района — центр и периферия яйца — связаны тонкими цитоплазматическими мостиками, а всё промежуточное пространство заполнено желтком.