Хромосомная теория наследственности. Сцепленное наследование.
Г. Мендель проследил наследование семи пар признаков у гороха. Многие исследователи, повторяя опыты Менделя, подтвердили открытые им законы. Было признано, что эти законы носят всеобщий характер. Однако в 1906 г. английские генетики В.Бэтсон и Р.Пеннет, проводя скрещивание растений душистого горошка и анализируя наследование формы пыльцы и окраски цветков, обнаружили, что эти признаки не дают независимого распределения в потомстве. Потомки всегда повторяли признаки родительских форм. Постепенно факты исключений из третьего закона Менделя накапливались. Стало ясно, что не для всех генов характерно независимое распределение в потомстве и свободное комбинирование. Каждый организм имеет огромное количество признаков, а число хромосом невелико. Следовательно, каждая хромосома несет не один ген, а целую группу генов, отвечающих за развитие разных признаков.
Убедительные доказательства того, что гены располагаются в хромосомах, были получены в 1910 г. американским генетиком, будущим лауреатом Нобелевской премии (1933 г.) Т. Морганом.
Многочисленные эксперименты Моргана и его сотрудников привели к ряду важнейших открытий, которые легли в основу хромосомной теории наследственности. Одно из положений этой теории можно сформулировать следующим образом: гены расположены в хромосомах в линейном порядке и занимают определенные участки — локусы, причем аллельные гены находятся в одинаковых локусах гомологичных хромосом. Совместное наследование генов, локализованных в одной хромосоме, известно под названием закона Моргана
Закон независимого наследования (третий закон Менделя) справедлив только в том случае, если неаллельные гены находятся в разных парах хромосом. Однако количество генов у живых организмов значительно больше числа хромосом. Например, у человека около 25 тыс. генов, а количество хромосом — 23 пары (2n = 46), у плодовой мушки дрозофилы приблизительно 14 тыс. генов и всего 4 пары хромосом (2n = 8). Следовательно, каждая хромосома содержит множество генов..
|
|
Гены, расположенные в одной хромосоме, образуют группу сцепления и наследуются вместе. Такое совместное наследование генов Т. Морган назвал сцепленным наследованием (в отличие от независимого). Гомологичные хромосомы каждой пары содержат гены, контролирующие одни и те же признаки, поэтому количество групп сцепления равно числу пар хромосом. Вам известно, что при независимом наследовании дигетерозиготная особь, например 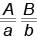 , образует четыре типа гамет в равном соотношении, т. е. по 25 %:
, образует четыре типа гамет в равном соотношении, т. е. по 25 %: 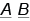 ,
, 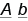 ,
, 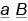 и
и  . Это обусловлено тем, что неаллельные гены находятся в разных парах хромосом. Если же они расположены в гомологичных хромосомах, следовало бы ожидать, что дигетерозигота
. Это обусловлено тем, что неаллельные гены находятся в разных парах хромосом. Если же они расположены в гомологичных хромосомах, следовало бы ожидать, что дигетерозигота 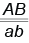 будет производить лишь два типа гамет: 50 %
будет производить лишь два типа гамет: 50 % 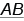 и 50 %
и 50 % 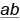 .
.
Однако Т. Морган обнаружил, что в большинстве случаев дигетерозиготные особи образуют не два типа гамет, а четыре. Кроме ожидаемых и , формируются также гаметы с новыми комбинациями генов: 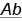 и
и 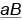 , но в меньшем процентном соотношении, чем при независимом наследовании. Рассмотрим один из опытов, в котором Т. Морган изучал наследование сцепленных генов у дрозофилы.
, но в меньшем процентном соотношении, чем при независимом наследовании. Рассмотрим один из опытов, в котором Т. Морган изучал наследование сцепленных генов у дрозофилы.
 Дрозофила обыкновенная, или плодовая, — вид насекомых отряда Двукрылые (рис. 34.1). Особи, как правило, имеют желто-коричневое брюшко с темными поперечными кольцами и глаза красного цвета. В природе эти мушки питаются опавшими фруктами, соком растений, гниющими растительными остатками. В XX в. дрозофила стала одним из самых распространенных объектов исследований в области генетики и биологии индивидуального развития. Удобство ее использования обусловлено рядом причин. Так, дрозофилы имеют множество отчетливо различимых альтернативных признаков, небольшие размеры (обычно около 2—2,5 мм), легко разводятся в лабораторных условиях и обладают высокой плодовитостью — от каждого скрещивания можно получить до нескольких сотен потомков. Важно также и то, что время развития особи от откладки яйца до выхода половозрелой мушки из куколки при 25 °C занимает всего 10 дней.
Дрозофила обыкновенная, или плодовая, — вид насекомых отряда Двукрылые (рис. 34.1). Особи, как правило, имеют желто-коричневое брюшко с темными поперечными кольцами и глаза красного цвета. В природе эти мушки питаются опавшими фруктами, соком растений, гниющими растительными остатками. В XX в. дрозофила стала одним из самых распространенных объектов исследований в области генетики и биологии индивидуального развития. Удобство ее использования обусловлено рядом причин. Так, дрозофилы имеют множество отчетливо различимых альтернативных признаков, небольшие размеры (обычно около 2—2,5 мм), легко разводятся в лабораторных условиях и обладают высокой плодовитостью — от каждого скрещивания можно получить до нескольких сотен потомков. Важно также и то, что время развития особи от откладки яйца до выхода половозрелой мушки из куколки при 25 °C занимает всего 10 дней.
|
|
Чистую линию дрозофил, имеющих серое тело и нормальные (длинные) крылья, скрестили с чистой линией, особи которой имели черное тело и зачаточные крылья (рис. 34.2). Полученные гибриды первого поколения в соответствии с первым законом Менделя были единообразными — серыми с нормально развитыми крыльями. Следовательно, у дрозофилы серое тело (А) полностью доминирует над черным (а), а нормальные крылья (В) — над зачаточными (b). Все гибриды первого поколения — дигетерозиготы.
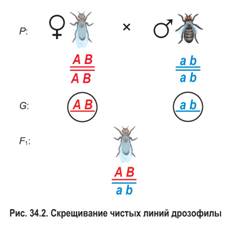
Затем было проведено анализирующее скрещивание (рис. 34.3). Дигетерозиготную самку из гибридного поколения скрестили с рецессивным дигомозиготным самцом (черное тело и зачаточные крылья). В потомстве было получено по 41,5 % особей с серым телом, нормальными крыльями и черным телом, зачаточными крыльями, а также по 8,5 % мух с серым телом, зачаточными крыльями и черным телом, нормальными крыльями.
|
|
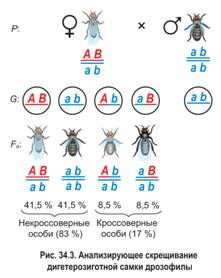
Если бы гены, определяющие цвет тела и развитие крыльев, находились в разных парах хромосом, соотношение фенотипических классов было бы равным — по 25 %, а числовое соотношение фенотипов не соответствует соотношению 1:1:1:1. Но этого не наблюдалось, значит, гены находятся в гомологичных хромосомах и наследуются сцепленно.
Несмотря на сцепление генов, самка производила не два, а четыре типа гамет. Однако гамет с исходными сочетаниями сцепленных генов формировалось намного больше ( и вместе составили 83 %), чем с новыми их комбинациями (сумма и равна 17 %).
Было выяснено, что причиной появления хромосом с новыми комбинациями родительских генов является кроссинговер. Этот процесс происходит в профазе I мейоза и представляет собой обмен соответствующими участками между гомологичными хромосомами. Таким образом, кроссинговер препятствует полному (абсолютному) сцеплению генов.
В зависимости от особенностей образования гамет, различают: кроссоверные гаметы — гаметы с хромосомами, претерпевшими кроссинговер; некроссоверные гаметы — гаметы с хромосомами, образованными без кроссинговера.
Соответственно этому различают: рекомбинантные (кроссоверные) особи — особи, возникшие с участием кроссоверных гамет; нерекомбинантные (некроссоверные) особи — особи, возникшие без участия кроссоверных гамет.
Гены в хромосомах имеют разную силу сцепления. Сцепление генов может быть: полным, если между генами, относящимися к одной группе сцепления, рекомбинация невозможна и неполным, если между генами, относящимися к одной группе сцепления, возможна рекомбинация. В рассмотренном эксперименте гаметы и являлись кроссоверными, а гаметы и — некроссоверными (см. рис. 34.3).
Т. Морган подвергал анализирующему скрещиванию не только дигетерозиготных самок дрозофилы, но и самцов, т. е. проводил реципрóкные скрещивания. Так называют пару скрещиваний, в одном из которых организмы с исследуемыми признаками (или признаком) используются в качестве материнских, а в другом — в качестве отцовских. Оказалось, что реципрокные анализирующие скрещивания дигетерозиготных особей дрозофилы дают разные результаты.
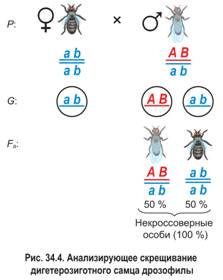
В потомстве дигетерозиготного самца наблюдалось лишь два фенотипических класса (рис. 34.4). Половину потомков составляли особи с серым телом и нормальными крыльями, вторую половину — особи с черным телом и зачаточными крыльями. Это свидетельствовало об отсутствии кроссинговера между сцепленными генами или, иначе говоря, о полном (абсолютном) сцеплении генов. Выяснилось, что у самцов дрозофилы в ходе мейоза действительно не происходит кроссинговер между гомологичными хромосомами. Впоследствии подобное явление было обнаружено у самцов некоторых других видов двукрылых, а также у самок тутового шелкопряда. Однако полное сцепление генов наблюдается в природе очень редко, как исключение из общего правила.
Кроссинговер между сцепленными генами происходит с определенной вероятностью (частотой). Для расчета частоты кроссинговера (rf, от англ. recombination frequency — частота рекомбинации) можно пользоваться следующей формулой:
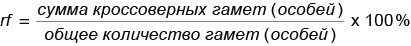 .
.
Таким образом, между генами А и В, контролирующими цвет тела и длину крыльев дрозофилы, кроссинговер происходит с частотой: rfAB = 17 %.
Расстояние между этими генами равно17 морганидам. А сила сцепления высчитывается по формуле: сила сцепления = 100% - % кроссоверных гамет. Сила сцепления между генами окраски тела и формы крыльев равна 100% - 17% = 83%.
Дальнейшие исследования, проведенные Т. Морганом и сотрудниками его лаборатории, показали, что частота кроссинговера пропорциональна расстоянию между генами, расположенными в одной хромосоме. Чем больше расстояние между сцепленными генами, тем чаще между ними происходит кроссинговер. И наоборот, чем ближе друг к другу расположены гены, тем меньше частота кроссинговера между ними.
В профазе I мейоза при конъюгации гомологичных хромосом взаимный обмен участками между хроматидами осуществляется произвольно, в любой их точке. Рассмотрим рисунок 34.5.
1. 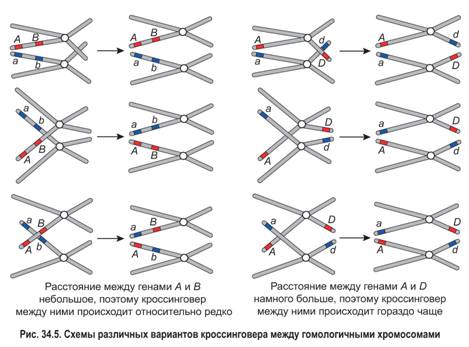
Гены А и В (или а и b) находятся сравнительно близко друг к другу. Вероятность того, что обмен произойдет именно на участке, разделяющем эти гены, невелика. Гены А и D (или а и d) располагаются на значительном расстоянии друг от друга. Поэтому вероятность того, что хроматиды совершат обмен в какой-либо точке между этими генами, намного выше. Значит, чем больше расстояние между генами, тем чаще они разделяются при кроссинговере.
Таким образом, частота кроссинговера позволяет судить о расстоянии между генами. В честь Т. Моргана единица измерения расстояния между генами получила название морганида или, что то же самое, сантиморган (сМ).
Морганида (сантиморган, сМ) — это генетическое расстояние, на котором кроссинговер происходит с вероятностью 1 %.
Кроссинговер играет очень важную биологическую роль. В результате этого процесса у потомков возникают новые комбинации родительских генов, что повышает генетическое разнообразие организмов и расширяет возможности их адаптации к различным условиям окружающей среды.
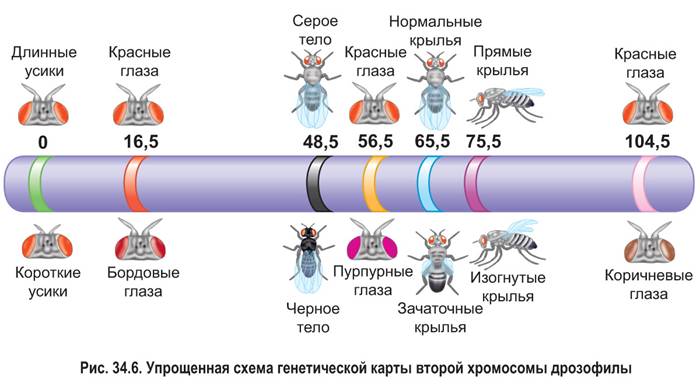
Генетические карты. Как показали эксперименты Т. Моргана, результаты анализирующего скрещивания дают возможность определять частоту кроссинговера между сцепленными генами и судить о расстоянии между ними (за исключением тех редких случаев, когда кроссинговер отсутствует). Использование данных, полученных от множества анализирующих скрещиваний с различными комбинациями сцепленных генов, позволяет ученым составлять так называемые генетические карты (рис. 34.6).
Генетическая карта хромосомы — это схема взаимного расположения генов, находящихся в одной группе сцепления, построенная с учетом расстояний между ними. Такие карты уже составлены для человека и многих других видов живых организмов, например для мыши, дрозофилы, гороха, пшеницы, кукурузы, томата, дрожжей и т. д.
.
Генетические карты находят широкое применение в научных исследованиях и практической деятельности человека. Знание генетических карт позволяет предсказывать характер наследования тех или иных признаков. Использование этих данных в селекции и генетической инженерии способствует планированию работы по получению организмов с определенными сочетаниями признаков. Сравнение генетических карт разных видов организмов дает возможность устанавливать степень их родства, судить о возможных путях эволюционных преобразований наследственного материала.