Гликоген является депо (запасом глюкозы). Гликоген образуется в печени и других тканях:
- Н2О
Ø  Глюкоза + АТФ Глюкоза-6-фосфат + АДФ
Глюкоза + АТФ Глюкоза-6-фосфат + АДФ
гексокиназа
(глюкокиназа при больших
концентрациях сахара)
Гексокиназа малоспецифична, катализирует эту реакцию и с другими гексозами (фруктозой, маннозой и т.д.)
Ø 
 Глюкозо-6-фосфат Глюкозо-1-фосфат
Глюкозо-6-фосфат Глюкозо-1-фосфат
фосфоглюко-мутаза
 Глюкозо-1-фосфат +УТФ УДФ - глюкоза + ПФ
Глюкозо-1-фосфат +УТФ УДФ - глюкоза + ПФ
УДФ – глюкозопирофосфорилаза
ПФ – соединение не стойкое и быстро расщепляется
Н2О
 ПФ 2Фн, поэтому реакция образования УДФ - глюкозы
ПФ 2Фн, поэтому реакция образования УДФ - глюкозы
пирофосфотаза
практически необратима.
Ø  (Глюкоза)n + УДФ/ глюкоза (Глюкоза)n+1 + УДФ
(Глюкоза)n + УДФ/ глюкоза (Глюкоза)n+1 + УДФ
(порция гликогена) а)при образовании a-1,4-глюкогидных
n – не менее четырёх связей работает фермент; гликогенсинтаза
б)при образовании a-1,6-гликозидных
связей работает фермент; амило-1,4-1,6-
гликозилтрансфераза.
Ветвление цепи гликогена повышает его растворимость и, благодаря созданию большого количества не восстанавливающих концевых остатков, которые являются местами действия гликоген-синтазы, увеличивает скорость синтеза и расщепления гликогена.
По мере необходимости гликоген может расщепляться (и в печени, и в других тканях). Различают два пути расщепления гликогена:
Ø Амилолитический
Таким путём расщепляются боковые цепи – это около 10% гликогена. В тканях есть a-амилазы (расщепляют срединные a-1,4-гликозидные связи) и a-амилазы (расщепляют концевые a-1,4- гликозидные связи). По сути, идёт гидролиз гликогена:
Н2О
 Гликоген Глюкоза
Гликоген Глюкоза
Амилаза
Полученную глюкозу, в какие бы процессы она потом ни вступала (синтез, распад), необходимо проактивизировать, т.е. получить фосфориокислый эфир глюкозы, т.е. фосфорилировать её:
 Глюкоза +АТФ Глюкоза-6-Фосфат + АДФ
Глюкоза +АТФ Глюкоза-6-Фосфат + АДФ
Как видно из приведённой реакции, мы затрачиваем при этом одну молекулу АТФ.
Ø Фосфоролитический (фосфоролиз):
Этим путём расщепляется до 90 % гликогена. Процесс выглядит по – другому:
1.  Гликоген + Н3РО3 Глюкозо-1-фосфат
Гликоген + Н3РО3 Глюкозо-1-фосфат
фосфорилаза
2.  Глюкозо-1-фосфат Глюкозо-6-фосфат
Глюкозо-1-фосфат Глюкозо-6-фосфат
фосфоглюкомутаза
Фосфорилаза расщепляет a-1,4- гликозидные связи, а a-1,6- гликозидные связи расщепляют a-1,6- гликозидазой. Но в реакции фосфоролиза участвует только активная форма – фосфорилаза «а», в то время, как в тканях вырабатывается и постоянно присутствует неактивная форма - фосфорилаза «b». Это впервые обнаружили в 1947 г. Дж. Корн, К. Корн. Превращение фосфорилазы «b» (неактивной) в фосфорилазу «а» (активную), происходит путём фосфорилирования. Фосфорилаза «b» представляет собой димер: каждая цепь содержит 841 АК, с молекулярной массой 100 000. При активации происходит фосфорилирование одного остатка серина (в 14 положении) в каждой субъединице.
 Фосфорилаза «b» + АТФ Фосфорилаза «а» + АДФ
Фосфорилаза «b» + АТФ Фосфорилаза «а» + АДФ
 | |||
| |||
сер – ОН сер – О – РО3Н2
Этот процесс катализирует фермент: киназа фосфорилазы. (Активизирующее влияние на фосфорилазу «b» оказывает и АМФ (его много образуется при повышенном потреблении АТФ и, следовательно, возникает большая потребность в АТФ). АМФ оказывает активирующее влияние на фосфорилазу «b» вследствие аллостерического эффекта – путём связывания с регуляторным центром на каждой из субъединиц). Но этот фермент тоже вырабатывается неактивным, и также активизируется путём фосфорилирования. Однако частично активизировать киназу фосфорилазы могут и Са++ в концентрации 10-7м. Этот механизм активации имеет для организма важное значение, т.к. мышечное сокращение запускается высвобождением Са++-ионов. Таким образом, ращепление гликогена и мышечное сокращение связаны преходящим увеличением содержания Са++ в цитоплазме.
Фосфорилированию же киназы фосфорилазы способствует фермент протеинкиназа, которая активна лишь при присутствии в цитоплазме циклической формы АМФ.
АМФ образуется в плазматической мембране из АТФ:
АТФ: Аденин-рибоза-Ф-Ф-Ф Аденин-рибоза- Ф + ПФ
Аденилатциклаза (цАМФ)
Аденилатциклаза - мембраносвязанный фермент. На его активность в значительной степени влияют специфические гормоны.
Мы видим, что синтез и расщепление гликогена происходит разными путями. Главные ферменты синтеза и распада гликогена – это гликогенсинтеза и фосфорилаза. Синтез и расщепление гликогена координировано регулируются таким образом, что когда активируется гликогенсинтаза, то фосфорилаза остаётся почти неактивной и наоборот. На обмен гликогена большое влияние оказывают специфические гормоны:
инсулин (вырабатывается в поджелудочной железе (b-клетки)-повышай способность печени и мышц синтезировать гликoген (интимный механизм инсулина пока выяснен не в достаточной степени),Значительно лучше изучен механизм действия адреналина и глюкогона.
адреналин вырабатывается мозговым веществом надпочечников и вы-раженно стимулирует распад гликогена в мышцах и, в меньшей мере, в печени. Печень более чувствительна к гликогону, который вырабатывается a –клетками поджелудочной железы (при низком содержании глюкозы в крови).
Эрл Сазерленд установил, что действие адреналина и глюкогена на обмен углеводов опосредуется циклическим АМФ. Адреналин и глюкогон не проникают в свои клетки-мишени, они связываются с плазматическими мембранами и стимулируют аденилатциклазу.
Таким образом,получается следующая схема:
Адреналин (глюкагон)
 |
Аденилатциклаза
 |
 АТФ цАМФ
АТФ цАМФ
 |
Протеинкиназа

 Киназа фосфорилазы
Киназа фосфорилазы
 Фосфорилаза Фосфорилаза
Фосфорилаза Фосфорилаза
«b» «а»
Фосфоролиз (распад гликогена)
Глюкозо-1-фосфат
Глюкозо-6-фосфат
Синтез гликогена тесно связан с его расщеплением. Активность гликоген-синтазы, подобно активности фосфорилазы, регулируется путем ковалентной модификации, т. е. фосфорилирования. В результате фосфорилирования гликоген-синтаза «а» (активная формула), превращается в неактивную форму-гликоген – синтазу «в».Фосфорилированная форма –гликоген-синтаза «в» требует для своей активности больших количеств глюкозо-6-фосфата.,тогда как дефосфорилированная «а» –форма активна и в присутствии и в отсутствии глюкозо-6-ффосфата. Таким образом,фосфорилирование оказывает противоположно направленное действие на активности гликоген-синтазы и гликоген-фосфорилазы. Так как фосфорилирование гликоген-синтазы «а» осущест-вляется протеинкиназой,то,когда протеин-киназа активна (см. схему, то-есть под действием адреналина и глюкогона), то она стимулирует выработку активной фосфорилазы «а» (через стАДНю киназы фосфорилазы) и неактивной гликоген-синтазы «в». Т. е. при повышенном распаде гликогена тормозится его синтез и наоборот.
Адреналин (глюкагон)
|
Аденилатциклаза
 |
 АТФ цАМФ
АТФ цАМФ

 Протеникиназа Гликогенсинтаза «b»
Протеникиназа Гликогенсинтаза «b»
фосфорилирование
 Cа++ Киназа фосфорилазы Гликогенсинтаза «а»
Cа++ Киназа фосфорилазы Гликогенсинтаза «а»
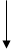 | |||
 | |||
 Фосфорилаза «b» Фосфорилаза «а»
Фосфорилаза «b» Фосфорилаза «а»
 | |||||
 | |||||
| |||||
УДФ-глюкоза
 | |||
| |||
Глюкоза-1-фосфат Глюкоза-1-фосфат
 |
Глюкоза-6-фосфат Глюкоза-6-фосфат
| |||
 | |||
СО2+ Н2О Глюкоза
Гликолиз.
Под гликолизом в общем смысле принято понимать разложение углеводов. В зависимости от конкретного организма и (или) условий его роста, гликолиз может идти по анаэробному пути (в этом случае он так и называется –анаэробный гликолиз).
И может идти по аэробному пути (соответственно – аэробный гликолиз). Для многих микроарганизмов, растущих в анаэробных условиях (в отсутствие кислорода) гликолиз является основным катаболическим путем для получения энергии из углеводных субстратов, приводя к образованию определенных конечных продуктов метаболизма, например, этанола, лактата, глицерина. Процесс такого типа часто называют брожением. У животных анаэробный гликолиз (с образованием лактата) обеспечивает энергией сокращение скелетных мышц в условиях ограниченного снабжения кислородом. В некоторых случиях гликолиз (аэробный гликолиз) идет по анаэробному пути в аэробных условиях т. е. конечным продуктом является лактат), хотя кислород и присутствует. Такие процес-сы идут, например, в эритроциттах, сетчатке, слизистой кишечника, в тканях плода (вскоре после рождения). Гликолиз –единственный анаэробный путь, дающий энергию. В аэробных условиях (в присутствии кислорода) реакции гликолиза составляют начальную форму разложения углеводов,связанную далее с циклом лимонной кислоты. В этом случае гликолиз останавливается на стАДНи образования ПВК. В аэробных условиях гликолиз и цикл Кребса совместно приводят к полному окислению гексозы до углекислого газа. Этот процесс сопровождается выделением больших количеств потенциально доступной метаболической энергии, в основном в виде высокоэнергетических продуктов НАДН и ФАДН2,которые, окисляясь в ходе дыхательной цепочки, дают энергию, необходимую для процесса ОФ, в результате которого образуется АТФ. Гликолиз –это первая до конца расшифрованная последовательность биохимических реакций, для этого потребовалось почти 50 лет (1890-1940). За это время были открыты фосфоэфиры, АТФ, НАД,выделены и охарактеризованы многие факторы и т.д.
Гликолиз протекает в цитозоле.Схема гликолиза:
АДФ

 СН2ОН СН2 ОРО3Н2
СН2ОН СН2 ОРО3Н2



 Н О Н АТФ Н О Н
Н О Н АТФ Н О Н



 Н Н
Н Н
 ОН Н Гексокиназа ОН Н Глюкозо-
ОН Н Гексокиназа ОН Н Глюкозо-
ОН ОН (глюкокиназа) ОН ОН фосфоизомераза
Н ОН Мg++ для всех киназ Н ОН
Глюкоза Глюкозо-6-фосфат
(Ингибитор гексокиназ: АТФ, Г-6-Ф, активатор: АДФ)

 Н2О3РОН2С О СН2ОН Н2О3РОН2С О СН2ОРО3Н2
Н2О3РОН2С О СН2ОН Н2О3РОН2С О СН2ОРО3Н2

 АДФ
АДФ
 Н ОН АТФ Н ОН
Н ОН АТФ Н ОН


 Н Н Фосфофруктокиназа Н Н альдолаза
Н Н Фосфофруктокиназа Н Н альдолаза
(тоже ингибируется)
ОН Н НАДН,АТФ ОН Н
Фруктозо-6-фосфат Фруктозо-1,6-дифосфат


 СН2ОРО3Н2 С = О НАД+ НАДН2
СН2ОРО3Н2 С = О НАД+ НАДН2
 Н +Н3РО4
Н +Н3РО4



 С=О Н – С – ОН
С=О Н – С – ОН
Триозофос- Глицеральдегидрофосфат-
СН2ОН фатизомераза СН2ОРО3Н2 -ДГ
Фосфодигидро- З-фосфоглицериновый отсюда начинается
ксиацетон альдегид окислительная стАДНя
 дифосфоглице- дифосфогли-
дифосфоглице- дифосфогли-
 ромутаза в эритроцитах церофосфатаза
ромутаза в эритроцитах церофосфатаза

 2,3 – ДФГ
2,3 – ДФГ 
СООН СООН

 С = О АДФ АТФ
С = О АДФ АТФ

 ОРО3Н2 Н – С – ОН Н – С – ОРО3Н2
ОРО3Н2 Н – С – ОН Н – С – ОРО3Н2
 Фосфоглицерат – Фосфоглицеро-
Фосфоглицерат – Фосфоглицеро-
Н – С – ОН киноза СН2ОРО3Н2 мутаза СН2ОН
3-фосфоглицерат 2-фосфоглицерат
СН2ОРО3Н2
1,3-ДФГ
 СООН СООН СООН
СООН СООН СООН
 - Н2О АДФ АТФ НАДН2 НАД+
- Н2О АДФ АТФ НАДН2 НАД+



 С – ОРО3Н2 пируватки- С=О Н – С – ОН
С – ОРО3Н2 пируватки- С=О Н – С – ОН
енолаза наза
СН2 СН3 СН3
Фосфоенол- ПВК Лактат
пируват
Очень важно образование на конечном этапе НАД+,который используется для окислительного фосфорилирования 3-фосфоглицеринового альдегида.
Лактат является конечным продуктом гликолиза. Т. к. лактат (как, впрочем, и пирува) достаточно легко проходит через плазматическую мембрану, он попадает из мышечной клетки (или эритроцитов) в кровь, с током крови попадает в печень, где окисляется в пируват, который в печени затем превращается в глюкозу. Таким образом образуется своего рода цикл. Эти превращения и составляют цикл Кори:


 Глюконеогенез Гликолиз
Глюконеогенез Гликолиз
| |||
| |||

 |  |
Кровь
| |||||
| |||||
 | |||||
Печень мышца
Вообще же в ходе гликолиза образуется 4 молекулы АТФ,а расходуется 2 АТФ. Таким образом, суммарный эффект окисления одной молекулы глюкозы в ходе гликолиза до лактата равен 2 АТФ. Утилизируется всего около 3% всей возможной энергии, которую можно было бы получать при окислении глюкозы, т.е. в сутки нужно было бы расщепить 27 кг глюкозы, чтобы обеспечить ортанизм энергией (если бы энергия получалась бы только путем гликолиза). Однако,гликолиз имеет место в клетках. Он нужен в мышцах для быстрого получения энергии. А, например, эритророциты только за счет гликолиза и могут получать энергию, т. к. у них нет митохондрий, и, следовательно, нет и окислительного фосфорилирования. Большинство простых сахаров можно расщепить гликолитическим путем. Все, что для этого необходимо – это превратить эти сахара в Г-1- Ф, Г-6-Ф, Ф-6-Ф. Это происходит в ходе взаимного превращения моносахаров.