Пирамидная система и периферические мотонейроны обеспечивают произвольные сокращения мышц. Каждый завершенный двигательный акт, каким бы простым он ни был, требует согласованного действия многих мышц. Качество движения зависит не только от вида и количества реализующих его мышц. Одни и те же мышцы участвуют в обеспечении различных движений. Вместе с тем одинаковое движение может производиться то медленнее, то быстрее, с меньшей или большей силой. Для выполнения движения необходимо подключение механизмов, регулирующих последовательность, силу и длительность мышечных сокращений и регламентирующих выбор необходимых мышц. Иными словами, двигательный акт формируется в результате последовательного, согласованного по силе и длительности включения отдельных нейронов корково-мышечного пути, отдающего распоряжения мышцам, и большого комплекса нервных структур вне пирамидной системы, которые объединяются в экстрапирамидную систему, действующую рефлекторно-автоматизированно.
Экстрапирамидная система включает клеточные группы коры больших полушарий (преимущественно лобных долей), подкорковые ганглии (хвостатое ядро - nucl, caudatus, скорлупа - putamen, латеральный и медиальный бледные шары - globus pallidus, субталамическое тело Льюиса), в стволе мозга черную субстанцию, красные ядра, пластинку крыши среднего мозга, ядра медиального продольного пучка (ядра Даркшевича), голубоватое место в мосту мозга, ретикулярную формацию с нисходящими и восходящими путями, мозжечок, γ-мотонейроны спинного мозга и др.
Между этими образованиями экстрапирамидной системы имеются многочисленные двухсторонние связи (замкнутые нейронные круги).
Произвольно выполняя любое действие, человек не задумывается о том, какую мышцу надо включить в нужный момент, не держит в сознательной памяти рабочую схему последовательности двигательного акта. Привычные движения производятся незаметно для внимания, смена одних мышечных сокращений другими автоматизированна. Эти двигательные автоматизмы способствуют наиболее экономному расходованию мышечной энергии в процессе выполнения движений. Новый, незнакомый двигательный акт энергетически всегда более расточителен, чем привычный, автоматизированный. Совершенствование качественной стороны движения с переводом их на автоматизированный наиболее экономичный режим обеспечивается деятельностью экстрапирамидной системы и в основном ее базальными ганглиями.
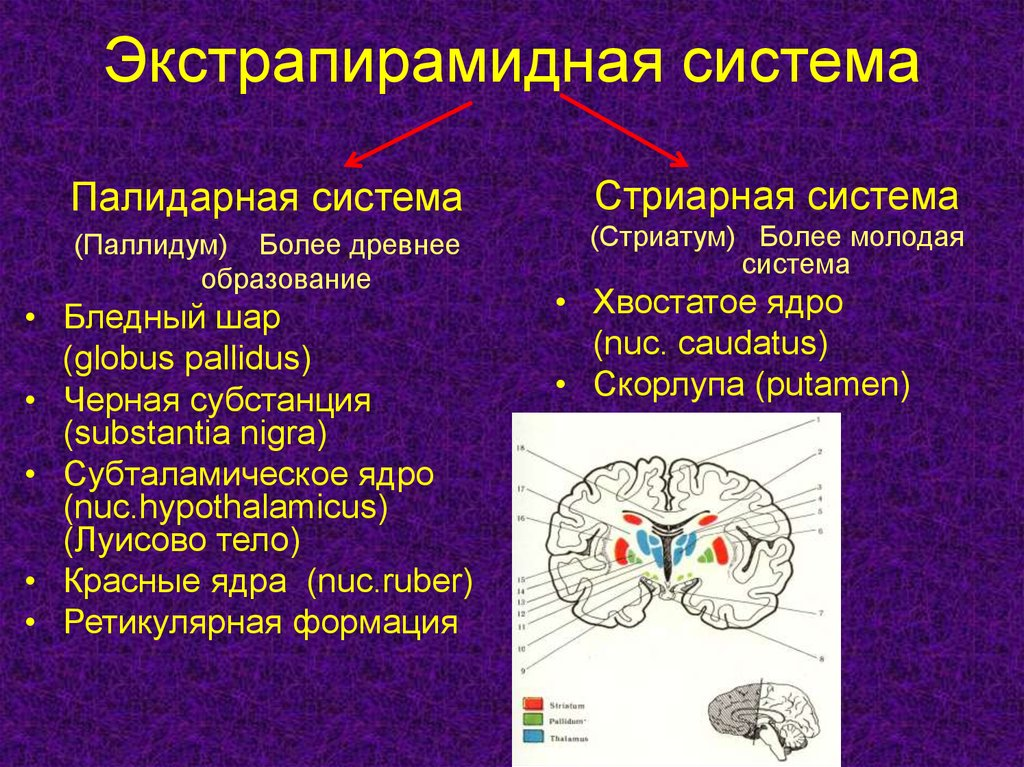
Морфологически и функционально стриопаллидарная система подразделяется на стриарную и паллидарную. Паллидарная система, филогенетически более старая, включает в себя латеральный и медиальный бледные шары, черное вещество, красное ядро, субталамическое ядро. В обоих бледных шарах содержится большое число нервных волокон, крупных нейронов в них относительно немного. Стриарная система филогенетически является “молодой” и включает хвостатое ядро и скорлупу с множеством мелких и крупных нейронов и сравнительно небольшим количеством нервных волокон. В стриарной системе имеется соматотопическое распределение: в передних отделах - голова, в средних - верхняя конечность и туловище, в задних отделах - нижняя конечность.
Паллидарная система у рыб и стриопаллидарная у птиц являются высшими двигательными центрами, определяющими поведение этих организмов. Стриопаллидарные аппараты обеспечивают диффузные движения тела, согласованную работу всей скелетной мускулатуры в процессе передвижения, плавания, полета и др. У высших животных и человека потребовалась более тонкая дифференцировка работы двигательных центров. В процессе эволюции возникла пирамидная система, которая подчинила себе стриопаллидарную систему.
В онтогенезе у человека миелинизация стриарных проводников заканчивается к 5-му месяцу жизни (раньше пирамидной системы), поэтому в первые месяцы жизни ребенка латеральный и медиальный бледные шары являются высшим двигательным центром. Моторика новорожденного носит явные “паллидарные” черты: излишество, своего рода щедрость движений, богатая мимика с улыбкой и др. С возрастом многие движения становятся все более привычными, автоматизированными, энергетически расчетливыми. Солидность и степенность взрослых являются своего рода торжеством стриопаллидарной системы над паллидарной.
При обучении целенаправленным движениям (включая и профессиональные, например игра на музыкальных инструментах, столярные, слесарные работы, вождение автомобиля и др.) можно выделить 2 фазы. Во время первой фазы (которую условно обозначают как паллидарную) движения чрезмерные, излишние по силе и длительности сокращения мышц. Вторая фаза (пирамидно-стриарная) заключается в постепенной оптимизации управления движениями. Они становятся энергетически рациональными и максимально эффективными и доводятся до автоматизма.
Экстрапирамидная система имеет многочисленные нейронные связи между своими образованиями, зрительным бугром и сегментарным двигательным аппаратом спинного мозга.
Все афферентные системы стриопаллидарной системы оканчиваются в полосатом теле. В эти системы входят пути от большинства областей коры мозга, от срединно расположенных ядер зрительного бугра (в частности, от медиального центрального и парафасцикулярного ядер таламуса), от компактной зоны черной субстанции и ядер срединного шва среднего мозга. От полосатого тела пути идут только в латеральный и медиальный бледные шары и ретикулярную зону черной субстанции, от которых, в свою очередь, начинаются основные эфферентные экстрапирамидные системы.
От медиального бледного шара аксоны идут к ядрам таламуса (медиальному центральному, вентролатеральному и переднему вентролатеральному ядрам). Отростки клеток этих ядер таламуса проецируются на двигательные и другие области коры лобной доли.
От черного вещества и бледного шара начинаются нисходящие системы, которые идут к ядрам покрышки среднего мозга и мозгового ствола, а отнихк двигательным нейронам спинного мозга в составе нигро-ретикулярно-спинно-мозгового и паллидо-ретикулярно-спинномозгового трактов.
От черного вещества имеются и восходящие пути к ядрам заднего таламуса и субталамическому ядру переднего таламуса. Нисходящие и восходящие эфферентные тракты черного вещества оказывают различное влияние на двигательную активность человека.
От клеток ядер ствола мозга начинаются аксоны, которые проходят в канатиках спинного мозга и заканчиваются синапсами с клетками передних рогов на разных уровнях. Ких числу относятся кроме упоминавшихся выше главных нигро- и паллидо-ретикулярно-спинномозговых путей преддверно-спинномозговой путь (tr. vestibulospinalis), оливо-спинномозговой (tr. olivospinalis), красноядерно-спинномозговой (tr. rubrospinalis, или путь Монакова), покрышечно-спинномозговой (tr. tectospinalis), медиальный продольный пучок (fasciculus longitudinalis medialis). Наиболее мощным оказывается ретикулярно-спинномозговой тракт. Он состоит из аксонов клеток ретикулярной формации и в спинном мозге проходит в переднем канатике (вентральная часть тракта) и в боковом канатике (медиальные и латеральные его отделы). Волокна покрышечно-спинномозгового пути на уровне продолговатого мозга образуют синапсы с клетками сетевидного образования, и этот пучок входит в состав ретикулярно-спинномозгового пути, вместе с которым спускается вниз в переднем канатике. То же надо сказать о проходящем в боковом канатике красноядерно-спинномозговом пучке. В пределах продолговатого мозга значительное число его волокон переходит в сетчатое вещество и спускается вниз в составе ретикулярно-спинномозгового тракта.
Волокна экстрапирамидной системы, как и пирамидной, также проходят вдоль всей цереброспинальной оси от коры до нижних отделов спинного мозга. Однако анатомо-гистологическое строение пирамидной и экстрапирамидной систем имеет существенное различие. Тела всех нейронов пирамидной системы сгруппированы в коре головного мозга. Схематически пирамидная система - это корковые нейроны с длинными аксонами, доходящими до разных сегментов спинного мозга. Экстрапирамидная система представляет собой длинную колонку клеток с большим количеством нервных волокон на протяжении всего головного и спинного мозга. Колонка эта местами резко увеличивается в объеме (подкорковые узлы), на некоторых уровнях образуется густое переплетение волокон с телами клеток (бледный шар, сетчатое вещество мозгового ствола и др.).
Открытию функционального значения экстрапирамидной системы способствовали клинические и особенно клинико-анатомические наблюдения; они выявили такие формы нарушения движения, которые нельзя объяснить ни поражением пирамидной системы, ни расстройством координации движений. Было описано много синдромов такого рода. При одном из них обращают на себя внимание замедленность и бедность движений, маскообразное лицо, вялая мимика, редкое мигание, общая скованность, отсутствие содружественных движений руками при ходьбе. Такая картина получила название гипокинеза (от греч. hypo - понижение, недостаточность; kinesis - движение). При другом типе заболевания развивается противоположное состояние. При осмотре больного бросаются в глаза автоматические насильственные движения. Их называют гиперкинезом (от греч. hyper - чрезмерное повышение и kinesis - движение). И при гипо- и при гиперкинезах наступает расстройство мышечного тонуса, заметно отличающееся от наблюдаемого при поражении пирамидной системы или периферического мотонейрона.
В 20-е годы текущего столетия была выдвинута концепция, что гипокинезы возникают вследствие поражения филогенетически более старого образования - бледного шара, а гиперкинезы возникают вследствие поражения хвостатого ядра и скорлупы. Полагали, что хвостатое ядро и скорлупа (новый стриатум) тормозят старый стриатум (бледный шар). Однако в последнее время такой механизм возникновения гипо- и гиперкинезов был отвергнут. Выяснено, что экстрапирамидные расстройства могут возникать при поражении и коры мозга, и его ствола.
В 40-60-х годах получены новые данные о функции той части экстрапирамидной системы, которую называют сетевидным образованием. Экспериментына животных показали, что раздражение этого образования электрическим током приводит к активации деятельности коры мозга. На электроэнцефалограмме видно, что медленная электрическая активность коры переходит в высокочастотную, низкоамплитудную (ресциядесинхронизации). Кроме того, в сетевидном образовании имеются участки, раздражение которых активирует деятельность и спинного мозга, приводит к усилению двигательных спинальных рефлексов. Это облегчающее действие на спинной мозг передается по ретикулярно-спинномозговому тракту. В составе сетевидного образования находятся зоны, раздражение которых вызывает торможение коры полушарий и спинного мозга. Было установлено, что импульсы, следующие по ретикулярно-спинномозговому тракту, достигают не только а-, но и γ-мотонейронов.
Таким образом, был выяснен конечный отрезок пути, по которому импульсы из экстрапирамидной системы поступают в скелетную мускулатуру. Афферентная часть дуги этого тонического рефлекса представлена волокнами глубокой мышечной чувствительности. На уровне мозгового ствола от этих проводников отходят коллатеральные волокна к сетевидному образованию. Возникающие в ней эфферентные импульсы могут проводиться вниз по ретикулоспинальному тракту. Восходящие афферентные импульсы из рецепторов мышечных веретен образуют 2 потока. Один из них проходит по классическому пути глубокой чувствительности, другой - по проводникам сетевидного образования. В результате активируется деятельность коры головного мозга, в частности - лобной доли, которая посылает импульсы не только к различным ганглиям и ядрам экстрапирамидной системы, но и непрерывно получает от них ответные сигналы (обратная афферентация). Образуются кольцевые системы, импульсы по которым идут не только от лобной доли к ганглиям и дальше на периферию, но и возвращаются через таламус обратно.
Сетевидное образование (fonnatio reticularis) является не только частью экстрапирамидной системы, но и частью вегетативно-висцеральной системы (лимбико-гипоталамо-ретикуляриый комплекс) в лобную долю. Принцип нейронного кольца, замыкающегося при помощи канала обратной связи, признают в настоящее время основным в организации деятельности центральной нервной системы.
В значительной мере прояснилась функция базальных ядер в связи с открытием роли церебральных трансмиттеров. Доказано, что в осуществлении регуляции двигательной функции большое значение имеют дофаминергаческие системы мозга. В полосатом теле находится более 80% от общего количества дофамина, а образующийся из дофамина норадреналин, также имеющий большое биологическое значение, содержится преимущественно в стволе мозга. На этом основании впервые было высказано предположение, что дофамин в мозге имеет собственное функциональное значение, связанное с экстрапирамидными образованиями, и отличное от его роли предшественника норадреналина, как думали раньше. Введение резерпина экспериментальному животному приводит к резкому уменьшению содержания дофамина в полосатом теле и к развитию акинетико-ригидного (паркинсоноподобного) синдрома. Последующее введение предшественника дофамина левовращающего изомера 3,4-дигидроксифенилаланина (1-дофа) устраняет данный синдром. Оказалось, что у больных с синдромом паркинсонизма резко снижено содержание дофамина в полосатом теле, и для лечения применяют 1-дофа.
Важным достижением последних лет являются открытие и гистологическая идентификация в мозге основных дофаминергических нейроналъных систем. Уже хорошо изучены 2 такие восходящие системы. Главную из них составляют аксоны меланинсодержащих нейронов компактной зоны черной субстанции, которые переходят в области покрышки и латеральном отделе гипоталамуса, затем входят в ножку мозга, далее идут через ретролентикулярный отдел внутренней капсулы, бледный шар и заканчиваются в полосатом теле характерными бусоподобными нервными терминалями. Эти терминали содержат большое количество дофамина, а также его метаболитов и синтезирующих ферментов. Дегенерация этого нигростриарного дофаминергического тракта с резким снижением синтеза и высвобождение дофамина из его терминалей в полосатом теле являются главными гистопатологическими и биохимическими признаками паркинсонизма. Это же лежит в основе характерного для такого заболевания клинического синдрома.
Второй восходящей дофаминергической системой является мезолимбический тракт. Он начинается от клеток интерпедункулярного ядра среднего мозга, расположенного медиально от черной субстанции, проходит сбоку от нигростриарного тракта и заканчивается в филогенетически более старых ядерных образованиях - nucl. accumbena, nucl. interstitialis striae tennmalis и tuberculum olfactorium, которые объединяются под названием лимбического полосатого тела, учитывая их связи с лимбической системой (гиппокамп, миндалина, обонятельная кора). Считается, что эта филогенетически древняя часть мозга связана со сложными поведенческими актами, обеспечивающими сохранность вида. От клеток лимбического полосатого тела аксоны идут в гипоталамус и кору лобной доли - структуры, которые участвуют в регуляции эмоциональных реакций и интеллектуальных функций; в частности, предполагается, что мезолимбический дофаминергический путь участвует в контроле за настроением и поведенческими реакциями. Кроме того, эта система нейронов контролирует начало двигательного акта и двигательные аффективные реакции (например, сопровождающие эмоции). Связывающим звеном между лимбической и экстрапирамидной двигательной системами является nucl. ассumbens. Это ядро расположено в вентромедиальной части передних отделов полосатого тела и имеет некоторые общие цитоархитектонические и биохимические характеристики. а также общие эфферентные проекции в черной субстанции и бледном шаре. У больных с паркинсонизмом, наряду с резким снижением содержания дофамина в полосатом теле, значительно уменьшается его содержание в nucl. accumbens и других лимбических образованиях. Очевидно, с этой биохимической патологией связаны акинезия и эмоциональные расстройства, характерные для паркинсонизма.
В полосатом теле выделено, кроме дофамина, еще 7 трансмиттеров: ацетилхолин, γ-аминомасляная кислота (ГАМК), норадреналин, серотонин, глутаминовая кислота и нейропептиды - субстанция Р и метэнкефалин. Предполагается участие в экстрапирамидной регуляции движений ангиотензина, который также находится и в стриопаллидонигральных системах. Однако функциональное значение нейропептидов в экстрапирамидной системе исследовано недостаточно. В развитии экстрапирамидной двигательной патологии существенное значение имеет, по-видимому, нарушение взаимодействия различных трансмиттеров с дофаминергическими системами мозга.
При электронной микроскопии в пределах полосатого тела выявлено 9 различных типов синапсов нейронов и определены их трансмиттеры. Более 90% клеток полосатого тела составляют шиловидные интернейроны. Именно с шипами дендритов интернейронов полосатого тела установлена синаптическая связь таких трактов, как нигростриарный дофаминергический, кортикостриарный глутаматергический и таламостриарный холинергический (трансмиттерацетилхолин). Основным трансмиттером между интернейронами является ацетилхолин, а в синапсах, образованных коллатералями аксонов проекционных клеток, разветвляющихся внутри полосатого тела, - ГАМК. Таким образом, на уровне интернейронального пула полосатого тела происходит конвергенция большинства афферентных трансмиттерных систем. Важным фактором является установление обратной связи между полосатым телом и черной субстанцией. Дофаминергический тракт меланинсодержащих клеток черной субстанции заканчивается на интернейронах полосатого тела, которые связаны посредством внутренних синаптических контактов с проекционными клетками. Or этих последних аксоны идут обратно к черной субстанции. Трансмиттером этого стрионигрального тракта является ГАМК.
Большие клетки передней части хвостатого ядра продуцируют субстанцию Р, которая транспортируется и осуществляет передачу импульсов на нейроны передней трети черной субстанции. На уровне интернейронов полосатого тела эта замкнутая система имеет контакты с глутаматергическим кортикостриарным путем и холинергическими рецепторами от медиального центрального и парафасцикулярного ядер таламуса. Результаты происходящей на интернейронах сложной переработки всей поступающей информации передаются через синаптические контакты с проекционными клетками на основные эфферентные системы, контролирующие двигательные функции. В этих механизмах дофамин оказывает подавляющее, а глутамат - возбуждающее действие на интернейроны полосатого тела, в то время как действие ацетилхолина зависит от его концентрации и адаптирует поступающие импульсы к необходимой в данный момент двигательной ситуации. Эта система усложняется синапсами (около 20%), образованными тремя основными афферентными трактами полосатого тела на больших проекционных клетках, в которых те же трансмиттеры оказывают противоположное действие: дофамин - возбуждающее, а глутамат - подавляющее.
Связь между nucl. accumbens и черной субстанцией, а также бледным шаром осуществляется трансмиттером ГАМК. Тормозное воздействие этого тракта по своим электрофизиологическим характеристикам отличается от влияния стриопаллидонигральных проекций.
Эти данные раскрывают интегративные механизмы полосатого тела, определяющие его основное функциональное значение, которое заключается в сосредоточении внимания и эмоций на одном наиболее важном в данный момент двигательном акте. Достигается это путем одновременного торможения всех других процессов, которые могут помешать его осуществлению. В выполнении этой роли большое значение придается ингибиторному ГАМК-ергическому стриопаллидонигральному пути, который регулирует активность различных по своей функциональной значимости эфферентных систем черной субстанции и бледного шара и отвечает за сосредоточение внимания к контралатеральной стороне. Предполагается, что при этом хвостатое ядро участвует в контроле за механизмами психической активности и в осуществлении психомоторных действий. Передача мотиваций к действию проводится по ГАМК-ергическому тракту от nucl. accumbens к черной субстанции. В то же время скорлупа регулирует основные двигательные механизмы через главные эфферентные экстрапирамидные системы, так называемые генераторы функций - нигральную систему, которая контролирует скорость движений, и паллидарную систему - основной, наиболее значимый локомоторный центр, регулирующий двигательные акты и внимание.
Следует отметить, что в отличие от черной субстанции не существует прямой обратной связи между globus pallidus и striatum. Эта связь является многоступенчатой и осуществляется через вентролатеральное ядро таламуса и нейроны премоторных областей коры.
Для осознания необходимости произвольного движения, его планирования и реализации в корковые нейроны поступает 2 потока импульсов. Один поток проходит через специфические корковые тракты различных видов чувствительности, а другой - через базальные ганглии и делает в них петлю. Проходящие по нему импульсы осуществляют подготовку мышц к движению и его осознание. В норме базальные ганглии обеспечивают автоматическую последовательность простых двигательных программ, необходимых для выполнения плана действия.
Менее исследовано, по сравнению с дофамином, значение в регуляции движений другого нейротрансмиттера - норадреналина. Основным норадренергическим центром в мозге является голубоватое место (locus coeruleus) в мосту мозга. Его нейроны образуют норадреналиновые нервные терминали во всех областях мозга, в том числе в коре больших полушарий и мозжечке, в гиппокампе. Таким образом, одно ядро через свои связи может оказывать влияние на многие структуры мозга. Иннервации такого характера не имеет никакая другая нейрональная система мозга. Обнаружены пути от голубоватого места до черной субстанции, по которому может осуществляться контроль активности дофаминергического нигростриарного тракта. У больных паркинсонизмом, наряду с патологией меланинсодержащих нейронов черного вещества, часто выявляются дегенеративные изменения в locus coeruleus со снижением содержания норадреналина в их системе.
В последние годы получены данные о модулирующем действии мозжечка на дофаминергические и норадренергические системы ствола мозга и базальных ганглиев. Известны анатомические эфферентные связи мозжечка с катехоламинергическими ядрами покрышки, черного вещества и полосатого тела, а также с вентролатеральным ядром таламуса. Экспериментально выявлены связи между полосатым телом и мозжечком. Повреждение мозжечка или его связей с катехоламинергическими ядрами может влиять на существующие экстрапирамидные нарушения, а также участвовать в развитии различных гиперкинезов.
Следует отметить, что в механизмах действия нейронных систем большую роль играет функциональное состояние катехоламиновых рецепторов, которые располагаются на мембранах постсинаптических и пресинаптических образований. Рецепторы постсинаптических мембран взаимодействуют с медиатором, выделяемым в синаптическую щель под влиянием нервных импульсов, и передают возбуждение на эффекторный орган или постсинаптический нейрон. В отличие от этого пресинаптические рецепторы располагаются на внешней поверхности мембран катехоламиновых терминалей. Их основное физиологическое значение заключается в модулированном высвобождении медиатора в синаптическую щель.
В норадренергических терминалах периферической нервной системы выявлено 3 вида пресинаптических рецепторов: тормозные β-рецепторы (они тормозят высвобождение норадреналина), облегчающие ^-рецепторы, и тормозные дофаминовые рецепторы. При стимуляции этих рецепторов снижается высвобождение норадреналина и затормаживается норадренергическая передача. Такой механизм лежит в основе развития гипотонии мышц, в частности при лечении 1-дофа больных с экстрапирамидными нарушениями движений.
В центральной нервной системе также обнаруживаются пресинаптические рецепторы. Выявлены они и на терминалях дофаминергических нейронов (тормозные дофаминовые ауторецепторы). В полосатом теле постсинаптические дофаминовые рецепторы локализуются на холинергических интернейронах. Активность нигростриарного дофаминергического пути контролируется через ГАМК-рецепторы, расположенные на дофаминергических нейронах черного вещества. Кроме того, на дофаминергических терминалях в полосатом теле обнаружены холинергические и опиатные рецепторы, которые имеют значение в модулированном высвобождении медиатора. Гиперчувствительность постсинаптических дофаминовых рецепторов в полосатом теле является важным патогенетическим фактором гиперкинезов.
В последние годы обнаружено существование двух видов дофаминовых рецепторов – D1 и D2, которые имеют разные фармакологические свойства. Воздействие на рецепторы D1 изменяет общую двигательную активность, тогда как стимуляция рецепторов D2 вызывает дискинезии.
Итак, функционально тесно связанная с пирамидными образованиями и системой координации экстрапирамидная система участвует в формировании мышечного тонуса и позы. Она как бы подготавливает скелетную мускулатуру в каждое данное мгновение воспринимать возбуждающие и тормозящие импульсы. Нарушение в одном из звеньев, регулирующих деятельность экстрапирамидной системы, может привести к появлению особой формы повышения мышечного тонуса - ригидности, а также к развитию гипо- или гиперкинеза.
При исследовании двигательной функции всегда приходится дифференцирование изучать компонент деятельности корково-мышечного пути и экстрапирамидной системы. Функция экстрапирамидной системы оценивается не по силе мышечного сокращения, а по качественной стороне движений. Обращают внимание на позу больного, выразительность речи, мимики и произвольных движений, включая ходьбу. Для оценки мышечного тонуса исследуются некоторые тесты.
Тест наклона головы: больной находится в положении лежа на спине, обследующий подкладывает свою кисть под затылочную область головы больного и наклоняет ее, а затем быстро переводит кисть ниже, на шею. У здорового человека происходит пассивное и быстрое разгибание в шейном отделе позвоночника. При экстрапирамидном повышении мышечного тонуса голова больного определенное время удерживается в приданном положении, затем плавно и медленно возвращается в исходное положение. Этот тест позволяет выявить ранние расстройства пластической ригидности. При выраженной ригидности мышц движения в шейном отделе позвоночника ограничены так же, как и при менингеальном синдроме ригидности затылочных мышц, и поэтому приведение подбородка к груди не удается. В отличие от менингита при экстрапирамидной ригидности нет болевого синдрома в затылочных мышцах при проведении этого теста.
Тест падения верхних конечностей: у стоящего больного обследующий поднимает его расслабленные верхние конечности в стороны несколько выше горизонтального уровня. Затем неожиданно быстро отпускает их и свои кисти перемещает на туловище обследуемого, чтобы определить время падения и силу удара верхних конечностей больного. При различии в тонусе мышц плечевого пояса справа, и слева выявится разница в скорости падения и силе удара.
Тест маятникового качания верхних конечностей: у здорового человека при ходьбе маятникообразные качания обеих верхних конечностей имеют одинаковую амплитуду, которая синхронна с шаговым движением контралатеральной нижней конечности. При экстрапирамидной ригидности отмечаются замедление и отставание в движении верхней конечности. Это особенно заметно при одностороннем поражении.
Тест маятникового качания нижних конечностей: больной сидит на высоком стуле (столе) со свободно свисающими нижними конечностями. Обследующий разгибает их в коленных суставах и свободно отпускает. У здорового человека при расслабленной мускулатуре нижних конечностей голени симметрично совершают несколько маятникообразных движений. На стороне повышения тонуса укорачивается время качания и уменьшается его амплитуда.
Тест Нойка-Ганева: при определении тонуса мышц верхней конечности при пассивных движениях в локтевом или лучезапястном суставе пациента просят активно поднять нижнюю конечность. Поднимание нижней конечности выявляет усиление пластической гипертонии мышц верхней конечности. В случае одностороннего поражения при активном движении нижней конечности на стороне повышения тонуса мышечный тонус на здоровой верхней конечности не повышается.
Тест Формана: в позе Ромберга с закрытыми глазами при экстрапирамидной ригидности тонус мышц верхних конечностей повышается, а в положении лежа на спине - понижается.
Тесты фиксации позы: при экстрапирамидной ригидности больной сохраняет на неопределенно долгое время любую позу, которую ему придают. Это проверяется следующими тестами:
а) тест разгибания в лучеэапястном суставе: врач производит полное разгибание в лучезапястном суставе и прекращает его, отпуская кисть; у больного с паркинсонизмом кисть находится в приданном положении, затем медленно сгибается в лучезапястном суставе;
б) стопный тест Вестфаля: резкое разгибание в голеностопном суставе (скользящим движением кисти врача по подошве) сопровождается застыванием стопы в приданном положении на какое-то время вследствие тонического напряжения мышц-разгибателей стопы (в основном - большеберцовой мышцы), затем стопа медленно опускается;
в) тест сгибания в коленном суставе: пациент находится в положении лежа на животе, врач сгибает расслабленные нижние конечности больного в коленных суставах под прямым углом и оставляетих в таком положении. При этом происходит рефлекторное сокращение мышц-сгибателей голени, нижняя конечность еще более сгибается в коленном суставе и длительное время удерживается в этом положении.
Оценка почерка: у больных с паркинсонизмом почерк становится мелким (микрография).
Основными синдромами экстрапирамидных поражений являются амиостатический синдром (паркинсонизм) и различные гиперкинезы.
Паркинсонизм. Для этого синдрома характерны монотонная тихая речь (брадилалия) и малая двигательная активность больного (олигокинезия). Лицо гипомимично, взгляд неподвижен, как будто устремлен в одну точку, мигание редкое, иногда оно отсутствует в течение нескольких минут, жестикуляция бедная. Туловище несколько наклонено вперед (поза просителя), верхние конечности согнуты в локтевых суставах, прижаты к туловищу. Отмечается наклонность к застыванию в какой-либо, даже неудобной, позе. Больной может лежать в постели с приподнятой над подушкой головой - симптом “воздушной подушки”. Активные движения совершаются очень медленно (брадикинезия). Затруднено начало двигательного акта - паркинсоническое “топтание на месте”. Ходит больной мелкими шажками, при ходьбе отсутствует обычная физиологическая синкинезия - содружественное движение верхних конечностей, они при ходьбе неподвижны (ахейрокинез). Иногда наблюдается еще один своеобразный симптом - пропульсия - больной на ходу начинает двигаться все быстрее и быстрее, не может остановиться и даже падает. Это объясняют тем, что перемещение центра тяжести не вызывает у больного реактивного сокращения мышц спины - “он как бы бежит за своим центром тяжести”. Если больному придать начальное движение (слегка толкнуть в грудь), он начинает двигаться назад (ретропульсия). Такое же вынужденное движение в сторону называют латеропульсией. Иногда наблюдаются “парадоксальные кинезии” (больные, которые в течение дня находятся в почти обездвиженном состоянии, в момент аффективных вспышек и эмоциональных напряжений могут прыгать, взбегать по лестнице, танцевать и др.)
Другой составной частью описываемого синдрома является мышечная ригидность - своеобразное сопротивление пассивным движениям. Ригидность мышц отличается от пирамидной спастичности тем, что она не только появляется в начальной фазе движения, но и удерживается во всех последующих фазах растяжения мышц. Конечность как бы застывает в той позе, которую ей придают. Такое состояние обозначают также как “пластический тонус”, “восковая гибкость” (flexibilitas cerea) и др.
Экстрапирамидная ригидность имеет и другие особенности: она наблюдается во многих мышечных группах (агонистах и антагонистах), но обычно преобладает в мышцах-сгибателях (“поза сгибателей”). Характерные для пирамидной спастичности явления “складного ножа” отсутствуют. Исследование пассивных движений может обнаружить прерывистость, как бы ступенчатость сопротивления мышц пассивным растяжениям (симптом “зубчатого колеса”).
Гипокинезия и ригидность могут наблюдаться изолированно, но к ним нередко присоединяется гиперкинез в виде тремора пальцев кисти, реже - нижних конечностей и подбородочной области. Этот ритмичный тремор пальцев кистей напоминает действие при счете монет или скатывании пилюль. Возникающая триада симптомов (гипокинез, ригидность, ритмичное дрожание) характерна для хронического заболевания, описанного в 1817 г. английским врачом J. Рагkinson и получившего название “дрожательный паралич”. Синдром, очень близкий по клинике к дрожательному параличу, был выявлен у больных в хронической стадии эпидемического энцефалита, с сосудистыми поражениями головного мозга, при некоторых экзогенных интоксикациях (соединениями марганца, аминазином, резерпином и др.). Он был назван паркинсонизмом. Наиболее выраженные гипертонус и тремор возникают при поражении черного вещества.
Из других симптомов при паркинсонизме характерны вегетативные расстройства (сальность лица, шелушение кожи, гиперсаливация и др.) и нарушения психоэмоционального тонуса. Последнее проявляется в виде снижения двигательной активности, аспонтанности в действиях. Иногда можно отметить описанную М. И. Аствацатуровым (1939) склонность больных к “приставанию” (акайрии) - навязчивому стремлению задавать одни и те же вопросы, повторно обращаться по незначительным поводам. Мышление у больных замедлено (брадипсихия).
Характерным для паркинсонизма является исчезновение всех его проявлений в период засыпания больного (понижается степень выраженности мышечного тонуса, прекращается тремор).
При длительном лечении паркинсонизма 1-дофа в виде побочных симптомов развиваются некоторые психопатологические и нейроэндокринные симптомы, что связано с нарушением функции дофаминсодержащих систем, таких как мезокортикальной (от ядра покрышки среднего мозга к коре лобной доли), тубероинфундибулярной (от нейронов агcuatus tuberculum infundibulum гипоталамуса к передней доле гипофиза).