Каждый организм подвергается многочисленным воздействиям. Эти воздействия можно подразделить на две большие группы: витальные — непосредственно влияющие на жизнеспособность (например, смертельная травма, причиненная хищником), и сигнальные — несущие информацию о внешней среде. Очевидно, что на многие сигналы необходимо реагировать, т.е. организм должен выработать определенный биологический ответ.
Общую схему формирования биологического ответа можно представить в виде трех последовательных этапов. На первом этапе происходит восприятие сигнала конкретным рецепторным белком, имеющим высокую специфичность к данному сигналу. Обнаружив сигнал, рецептор изменяет конформацию и тем самым извещает организм о наличии воздействия. Воспринятый сигнал необходимо преобразовать и передать в преобразованном виде на соответствующие структуры. Данный этап называют трансдукцией сигнала. Как правило, его осуществляют специальные белки (белки-посредники), а также различные вспомогательные молекулы и ионы. Существуют разные механизмы трансдукции (за счет либо синтеза, либо активации белков-посредников), но в конечном итоге сигнал достигает необходимых структур и приводит к запуску третьего, последнего этапа, т.е. к непосредственной реализации биологического ответа.
Различают 2 типа биологических ответов в зависимости от скорости формирования:
В частности, процессы каскадной активации белков могут осуществляться в организме очень быстро (иногда всего лишь за тысячные доли секунды) и поэтому обусловливают быстрые биологические ответы. Между тем каскады с использованием транскрипционных факторов требуют достаточно длительных промежутков времени (нескольких минут, часов или даже дней) и в итоге приводят к медленным биологическим ответам.
Быстрые биологические ответы реализуются за счет нервной системы. Гуморальная система – система медленных биологических ответов.
Молекулярные механизмы функционирования нервной системы тесно связаны с формированием и проведением нервных импульсов.
Исходной причиной, лежащей в основе формирования импульса, является восприятие нейроном определенного сигнала. Эту функцию выполняют молекулы соответствующего рецептора, расположенные, как правило, на наружной мембране нейрона. Обнаружив сигнал, рецепторы изменяют свою конформацию и тем самым воздействуют на соседние ионные каналы, заставляя их перейти из закрытого состояния в открытое.
Механизмы такого воздействия могут быть разными. В некоторых случаях (например, в чувствительных нейронах, обеспечивающих восприятие тепловых или механических сигналов) сами рецепторы одновременно являются и ионными каналами, поэтому активация данных рецепторных белков напрямую приводит к значительной деполяризации мембраны и в результате — практически к мгновенному формированию импульса. Такие рецепторы принято называть ионотропными. Но чаще всего между рецепторами и ионными каналами функционируют достаточно сложные каскады посредников.
Наиболее распространенные варианты подобных каскадов:
|
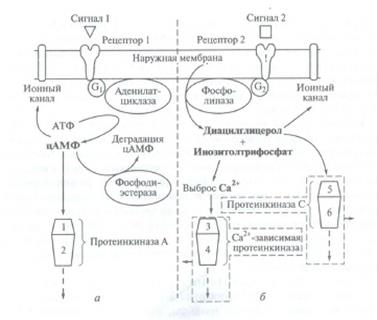
Несмотря на многочисленные нюансы, эти каскады организованы по сходному молекулярному принципу. В них участвуют следующие «наборы» компонентов:
• рецепторы, насквозь пронизывающие наружную мембрану;
• G-белки, расположенные на внутренней стороне мембраны и активируемые в результате изменения конформации рецепторов;
• фермент, активность которого регулируется G-белками. В одном варианте каскада это аденилатциклаза, в другом — фосфолипаза С или D;
• вторичные мессенджеры, т.е. небольшие регуляторные молекулы, служащие внутриклеточными переносчиками информации о сигнале. Они способствуют дальнейшей передаче и амплификации сигнала и характеризуются следующими свойствами: имеют небольшую молекулярную массу и с высокой скоростью диффундируют в цитоплазме; быстро расщепляются и быстро удаляются из цитоплазмы. К ним относится циклический аденозинмонофосфат (сокращенно — цАМФ), синтез которого обеспечивается аденилатциклазой из АТФ, находящихся в цитоплазме, а также инозитолтрифосфат и диацилглицерол, образуемые за счет активности фосфолипазы из молекулярных компонентов наружной мембраны. Появление цАМФ в цитоплазме должно быть импульсным, поэтому излишки данного вторичного мессенджера уничтожаются специальным ферментом — фосфодиэстеразой, активизируемой Са2+-зависимой протеинкиназой Нужно отметить, что появление в клетке инозитолтрифосфата приводит к выбросу Са2+ из эндоплазматической сети, и эти ионы, быстро диффундируя по всей цитоплазме, тоже становятся важными вторичными мессенджерами;
• белки, активируемые под действием вторичных мессенджеров. Это особые ионные каналы, обеспечивающие начальную стадию деполяризации мембраны. Кроме того, это определенные протеинкиназы, активно участвующие в регуляции последующих процессов за счет фосфорилирования различных мембранных белков.
Активация одной единственной молекулы рецептора приводит к едва ощутимому изменению потенциала. Между тем для того, чтобы открылись натриевые каналы (а без этого импульс принципиально не может сформироваться), необходима довольно существенная деполяризация мембраны: как правило, не менее чем на 20 мВ. И если воспринятый сигнал недостаточно интенсивен, т.е. активирует слишком мало молекул рецептора, возникшее изменение не достигает необходимого порога и быстро компенсируется различными ионными насосами. В результате мембрана возвращается к первоначальному состоянию, и выработки нервного импульса не происходит.
Казалось бы, данный принцип функционирования нейронов (он получил название «все или ничего») в корне противоречит тому общеизвестному факту, что организм не только распознает наличие сигналов, но и способен оценивать их интенсивность. Однако в действительности никакого противоречия нет. Взаимодействие между сигналом и молекулами рецептора, как и любые другие молекулярные взаимодействия, строго подчиняется статистическим (т.е. вероятностным) закономерностям. Это значит, что при действии сигнала фиксированной силы существует определенная вероятность формирования импульса. Чем мощнее сигнал, тем эта вероятность выше. Соответственно, чем выше интенсивность воспринимаемого сигнала, тем чаще нейрон вырабатывает нервные импульсы. Именно это и служит оценкой интенсивности сигнала.