У эукариотических организмов механизм регуляции транскрипции гораздо более сложен. В результате клонирования и секвенирования генов эукариот обнаружены специфические последовательности, принимающие участие в транскрипции и трансляции.
Для эукариотической клетки характерно:
1. Наличие интронов и экзонов в молекуле ДНК.
2. Созревание и-РНК - вырезание интронов и сшивка экзонов.
3. Наличие регуляторных элементов, регулирующих транскрипцию, таких как: а) промоторы - 3 вида, на каждый из которых садится специфическая полимераза. РНК-полимераза I реплицирует рибосомные гены, РНК-полимераза II - структурные гены белков, РНК-полимераза III - гены, кодирующие небольшие РНК. Промоторы РНК-полимеразы I и РНК-полимеразы II находятся перед участком инициации транскрипции, промотор РНК-полимеразы III - в рамках структурного гена; б) модуляторы - последовательности ДНК, усиливающие уровень транскрипции; в) (энхансеры) усилители - последовательности, усиливающие уровень транскрипции и действующие независимо от своего положения относительно кодирующей части гена и состояния начальной точки синтеза РНК; г) терминаторы - специфические последовательности, прекращающие и трансляцию, и транскрипцию.
Эти последовательности по своей первичной структуре и расположению относительно инициирующего кодона отличаются от прокариотических, и бактериальная РНК-полимераза их не "узнает". Таким образом, для экспрессии эукариотических генов в клетках прокариот нужно, чтобы гены находились под контролем прокариотических регуляторных элементов. Это обстоятельство необходимо учитывать при конструировании векторов для экспрессии.
Структурные гены, обеспечивающие жизнедеятельность эукариотической клетки, обычно транскрибируется в большинстве активно функционирующих клеток. В то же время, специфические гены, уникальные тех или иных тканей или органов, транскрибируются и транслируются только в определенных клетках. Например, гены, кодирующие α- и β-субъединицы гемоглобина взрослого человека, экспрессируются исключительно в клетках - предшественниках эритроцитов. Число разных мРНК, специфичных для разных клеток, варьирует от единиц до десятков.
Способность клеток включать (активировать) или выключать (ингибировать) структурные гены крайне важна для поддержания клеточной специфичности и экономного расходования энергетических ресурсов. Отсюда и многообразие факторов транскрипции, имеющих белковую природу. Многие из них связываются непосредственно с нуклеотидной последовательностью длиной менее 10 п.н., называемой по-разному: боксом, модулем, элементом инициации, регуляторным элементом. В отличие от прокариот у эукариот опероны в большинстве своем отсутствуют, т. е. каждый эукариотический структурный ген имеет свой собственный набор регуляторных элементов. Существенную роль в регуляции транскрипции у эукариот, помимо опосредованной взаимодействием между ДНК и белками, играют также белок-белковые взаимодействия.
Несмотря на индивидуальность набора регуляторных элементов у структурных генов эукариот, каждый из них имеет промоторный участок (ТАТА-бокс, или бокс Хогнесса) из восьми нуклеотидов, включающий последовательность TATA; последовательность ССААТ (САТ-бокс); участок из повторяющихся динуклеотидов GC (GC-бокс). Эти элементы находятся на расстоянии 25, 75 и 90 п.н. от сайта инициации соответственно:
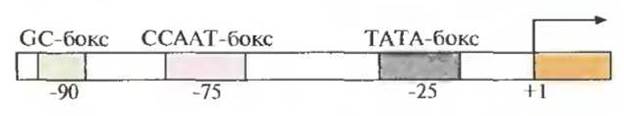
Регуляторные элементы структурных генов эукариот. Отрицательное значение показывает, что эти элементы находятся в молекуле ДНК слева от сайта инициации транскрипции, обозначенного +1. Стрелка — направление транскрипции (по Глик Б., Пастернак, Дж., 2002)
Транскрипция структурного гена эукариот начинается со связывания с ТАТА-боксом фактора транскрипции, который представляет собой комплекс по крайней мере из 14 белков. Затем с ним и участками ДНК, примыкающими к ТАТА-боксу, связываются другие факторы транскрипции, и, наконец, со всем этим транскрипционным комплексом связывается РНК-полимераза II. Затем при участии дополнительных факторов происходит инициация транскрипции в точке +1. Если последовательность TATA отсутствует или существенно изменена, то транскрипция структурного гена становится невозможной.
9. В конце 60-х годов работами американских ученых Р. Бриттена, Э. Дэвидсона и других была открыта фундаментальная особенность молекулярной структуры генома эукариот – нуклеотидные последовательности разной степени повторяемости. Это открытие было сделано с помощью молекулярно-биологического метода изучения кинетики ренатурации денатурированной ДНК. Различают следующие фракции в геноме эукариот.
1.Уникальные, т.е. последовательности, представленные в одном экземпляре или немногими копиями. Как правило, это цистроны – структурные гены, кодирующие белки.
2.Низкочастотные повторы – последовательности, повторяющиеся десятки раз.
3.Промежуточные, или среднечастотные, повторы – последовательности, повторяющиеся сотни и тысячи раз. К ним относятся гены рРНК (у человека 200 на гаплоидный набор, у мыши – 100, у кошки – 1000, у рыб и цветковых растений – тысячи), тРНК, гены рибосомных белков и белков-гистонов.
4. Высокочастотные повторы, число которых достигает 10 миллионов (на геном). Это короткие (~ 10 пн) не кодирующие последовательности, которые входят в состав прицентромерногогетерохроматина.
У эукариот объем наследственного материала значительно больше. В отличие от прокариот в эукариотических клетках одновременно активно транскрибируется от 1 до 10% ДНК. Состав транскрибируемых последовательностей и их количество зависят от типа клетки и стадии онтогенеза. Значительная часть нуклеотидных последовательностей у эукариот не транскрибируется вообще — молчащая ДНК.
Большой объем наследственного материала эукариот объясняется существованием в нем помимо уникальных также умеренно и высоко повторяющихся последовательностей. Эти высоко повторяющиеся последовательности ДНК располагаются в основном в гетерохроматине, окружающем центромерные участки. Они не транскрибируются. Характеризуя наследственный материал прокариотической клетки в целом, необходимо отметить, что он заключен не только в нуклеоиде, но также присутствует в цитоплазме в виде небольших кольцевых фрагментов ДНК —плазмид.
Плазмиды — это широко распространенные в живых клетках внехромосомные генетические элементы, способные существовать и размножаться в клетке автономно от геномной ДНК. Описаны плазмиды, которые реплицируются не автономно, а только в составе геномной ДНК, в которую они включаются в определенных участках. В этом случае их называют эписомами.
В прокариотических (бактериальных) клетках обнаружены плазмиды, которые несут наследственный материал, определяющий такие свойства, как способность бактерий к конъюгации, а также их устойчивость к некоторым лекарственным веществам.
В эукариотических клетках внехромосомная ДНК представлена генетическим аппаратом органелл — митохондрий и пластид, а также нуклеотидными последовательностями, не являющимися жизненно необходимыми для клетки (вирусоподобными частицами). Наследственный материал органелл находится в их матриксе в виде нескольких копий кольцевых молекул ДНК, не связанных с гистонами. В митохондриях, например содержится от 2 до 10 копий мтДНК.
Внехромосомная ДНК составляет лишь небольшую часть наследственного материала эукариотической клетки.