Поведенческие проявления тревожности у животных и тревожные расстройства у людей, в первую очередь, определяются состоянием центральной нервной системы. Поэтому для оценки роли опиоидной системы в нейрохимических механизмах тревоги было исследовано состояние центральных ОР и активность эндогенных лигандов этих рецепторов и ферментов деградации опиоидных пептидов в мозгу животных с различными поведенческими характеристиками.
На первом этапе исследования была обнаружена взаимосвязь между уровнем тревожности крыс при тестировании их в «челночной камере» и вытесняющей активностью эндогенных опиоидов в гиппокампе, которую определяли радиолигандным методом, используя лей-энкефалин для построения калибровочной кривой. Было показано, что процесс тестирования в «челночной камере» в несколько раз повышает уровень эндогенных опиоидов в гиппокампе крыс. Причем максимальный выброс опиоидов происходит в гиппокампе тех животных, которые не переходят в безопасный отсек «челночной камеры» после условного сигнала (пассивная стратегия избегания) (рис. 1).
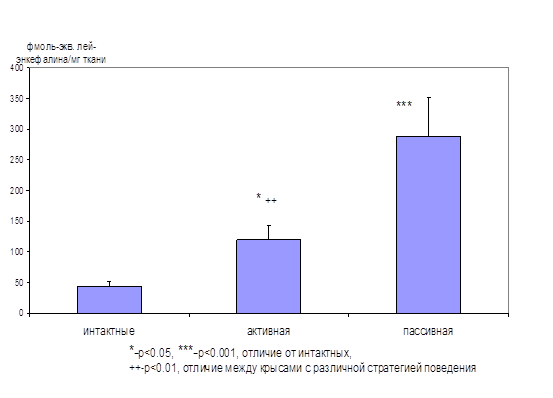
Рисунок. 1. Активность эндогенных лигандов ОР в гиппокампе крыс с различной стратегией поведения в «челночной камере»
Активная стратегия (19 крыс) – более 70% переходов в ответ на условный звуковой сигнал, пассивная (8 крыс) – остальные тестированные животные. Интактные животные (12) не были тестированы в «челночной камере». По оси ординат – способность уксуснокислых экстрактов из гиппокампов крыс вытеснять 3Н-ДАДЛЭ из ОР, нормированная в соответствии с калибровочной кривой, построенной с использованием лей-энкефалина.
Учитывая важную роль гиппокампа в регуляции эмоциональной сферы, но не болевой чувствительности, обнаруженные отличия скорее связаны с эмоциональным состоянием животных, уровнем их тревожности, чем с ноцицепцией. Отсутствие перехода на условный сигнал можно интерпретировать как сниженной способностью запоминать информацию об условном сигнале, так и пониженной тревожностью крыс. Результаты данного исследования дают основание утверждать, что противотревожную роль в этих условиях выполняют эндогенные опиоиды. Действительно, уровень опиоидов в гиппокампе крыс с пассивной стратегией поведения в 7 раз выше, чем у интактных животных (рис. 1), что, по-видимому, позволяет им преодолевать страх. Поэтому они не пытаются избегать боли и остаются в камере после условного сигнала. Косвенным подтверждением этому являются данные о том, что введение даларгина крысам приводит к изменению активной стратегии избегания на пассивную (Зозуля и соавт., 1996). Подобные эффекты обнаружены и при центральном введении селективного агониста δ1‑ОР (Ukai et al., 1997) и β-эндорфина (Flood et al., 1992).
Таким образом, у крыс с пассивной стратегией поведения в тесте «челночная камера», которую можно интерпретировать как относительно низкую тревожность, наблюдается значительно более высокий уровень эндогенных опиоидов в гиппокампе по сравнению с интактными животными и крысами с более высокой тревожностью. То есть, в условиях стресса в различных отделах мозга происходит усиление синтеза и секреции эндогенных опиоидов, которые оказывают анксиолитическое действие. В условиях хронического стресса, когда истощение запасов эндогенных опиоидов приводит к усилению поведенческих проявлений тревожности, становится более выраженным анксиолитическое действие их синтетических аналогов (табл. 1, Зозуля и соавт., 1999, Narita et al., 2006).
Для оценки состояния центральных ОР было проведено радиорецепторное определение констант связывания синтетического аналога лей-энкефалина Н-ДАДЛЭ с мембранной фракцией обонятельных луковиц и стриатума крыс. Стриатум был выбран как основной отдел мозга, отвечающий за двигательную активность, которая весьма существенна при тестировании поведения животных большинством использованных методов. Обонятельные луковицы, через которые поступает большая часть информации в мозг животных, особенно грызунов, являются одним из эмоциогенных отделов мозга.
Как было показано выше, при внутрибрюшинном введении агонист ОР даларгин оказывает анксиолитическое действие лишь в условиях стресса и не влияет на интактных животных. Поэтому в данном исследовании проводилось сравнение констант связывания ОР интактных и стрессированых животных. В качестве стрессорного воздействия были выбраны те же условия, что и для поведенческого тестирования, т.е. питьевая депривация крыс при пищевом режиме ad libitum. Перед началом стрессорного воздействия крысы были протестированы в ATS и в классическом варианте ОП, а по его окончании – в ПКЛ.
В нашем исследовании оказалось, что константы связывания ОР в стриатуме коррелируют с двигательной активностью крыс как в ATS, так и в ОП. Аффинность ОР стриатума к ДАДЛЭ у низкоактивных (НА) крыс выше, чем у высокоактивных (ВА) (табл. 3). Это соотношение отмечено как у интактных крыс, так и у крыс, подвергавшихся водной депривации. Cчитается, что Кд рецепторов может менять свои значения как из-за изменения микроокружения рецепторов (ионного состава среды, наличия оккупированных лигандами рецепторов других типов, липидного состава и состояния мембраны), так и по причине наличия нескольких типов или подтипов рецепторов, с которыми может взаимодействовать исследуемый лиганд. В нашем случае РРА проводится in vitro, поэтому причинами изменения Кд могли стать только изменение состояния мембраны или наличие нескольких подтипов ОР: μ-ОР, d1‑ОР и d2‑ОР.
Таблица 3. Константы связывания 3Н-ДАДЛЭ с центральными ОР у крыс с высокой (ВА) и низкой (НА) исходной двигательной активностью в ATS
| группы животных | Обонятельные луковицы | Стриатум | |||
| Кд (нМ) | Вмах (фмоль/мг белка) | Кд (нМ) | Вмах (фмоль/мг белка) | ||
| интактные | все | 2.9±0.2 (14) | 182±22 (14) | 2.4±0.2 (12) | 96±10 * (12) |
| ВА | 2.6±0.2 (8) | 179±33 (8) | 2.9±0.1## (7) | 114±27#* (7) | |
| НА | 3.0±0.3 (6) | 185±12 (6) | 2.0±0.3* (5) | 63±12** (5) | |
| стресс | все | 2.9±0.4 (9) | 170±27 (9) | 2.7±0.1 (14) | 100±7 (14) |
| ВА | 2.1±0.5## (4) | 152±14 (4) | 3.0±0.1#* (6) | 105±16 (6) | |
| НА | 3.8±0.3 (5) | 226±54 (5) | 2.4±0.2* (8) | 94±8* (8) |
* – p<0.05, ** – p<0.01 – отличие констант связывания в различных отделах мозга,
# – p<0.05, ## – p<0.01 – отличие констант связывания ОР у НА и ВА животных
Приведены средние значения ± ошибка среднего.
По литературным данным сродство ДАДЛЭ к δ-ОР в 2 раза выше, чем к μ-ОР (Kd=1.5 и 3.2 нМ, соответственно, при тестировании методом вытеснения селективных лигандов из δ- и μ-ОР мозга крысы, Leslie, 1987). То же соотношение Кд при несколько других абсолютных значениях получено на трансгенных клетках, стабильно экспрессирующих эти рецепторы (Minami, Saton, 1995, George et al., 2000). Судя по значениям Кд, полученным в нашем исследовании (табл. 3), в стриатуме ВА крыс доминируют μ-ОР, а у НА δ-ОР вносят свой вклад в соотношение ОР. Полученные данные соответствуют представлению о том, что ОР стриатума принимают участие в регуляции локомоторной активности путем воздействия на дофаминергическую трансмиссию (Jang et al., 2001, Schad et al., 2002). Причем как на фармакологическом, так и на молекулярно-генетическом уровнях показано, что активация μ-, но не δ-ОР приводит к повышению локомоции (Jang et al., 2001, Gaveriaux-Ruff, Kieffer, 2002).
Анализ взаимосвязи состояния центральных ОР с уровнем тревожности в ПКЛ показал, что до стресса водной депривации у менее тревожных крыс (количество выходов в открытые рукава ПКЛ больше, чем среднее по выборке, >2), по сравнению с более тревожными, повышена плотность ОР как в стриатуме, так и в обонятельных луковицах (табл. 4). Это косвенно свидетельствует о том, что в норме ОР δ- и μ-типов опосредуют «противотревожные» функции эндогенных опиоидов.
Таблица 4. Константы связывания 3Н-ДАДЛЭ с центральными ОР крыс с различным уровнем тревожности в ПКЛ
| Число выходов в откр. рукава | Обонятельные луковицы | Стриатум | |||
| Кд (нМ) | Вмах (фмоль/мг белка) | Кд (нМ) | Вмах (фмоль/мг белка) | ||
| ин- такт- ные | 2.6±0.3 (3) | 180±6## (3) | 2.3±0.3 (3) | 67±12#** (3) | |
| 1–2 | 2.6±0.1# (6) | 178±28 (6) | 2.4±0.3 (4) | 81±9#* (4) | |
| 3–5 | 3.6±0.4 (3) | 261±15 (3) | 2.6±0.2 (4) | 132±12* (4) | |
| стресс | 3.8±0.1# (4) | 203±49 (4) | 2.5±0.5* (4) | 88±15* (4) | |
| 1–2 | 2.6±0.5 (5) | 147±16 (5) | 2.7±0.5 (10) | 103±27 (10) |
* – p<0.05, ** – p<0.01 – отличие констант связывания в различных отделах мозга,
#,## – p<0.05, 0.01 – отличие от низкотревожных (по соответствующей группе) животных
Приведены средние значения ± ошибка среднего.
В результате использованного типа стрессорного воздействия происходит выраженное увеличение тревожности крыс, на фоне которого, как показано нами ранее, проявляется анксиолитический эффект даларгина, при этом на интактных животных препарат не влияет (табл. 1). По-видимому, длительный выброс эндогенных опиоидов в результате хронического стресса приводит к истощению их запасов. Даларгин в этих условиях оказывает анксиолитическое действие по компенсаторному механизму.
С другой стороны, известно, что в результате длительного контакта с агонистами может происходить десенситизация рецепторов. В частности, выброс эндогенных опиоидов при хроническом стрессе приводит к десенситизации ОР, причем δ-ОР более подвержены этому процессу, чем μ-ОР (Gaudriault et al., 1997, Kim et al., 2003, Narita et al., 2006). В нашем исследовании радиолигандный анализ не выявил существенных изменений констант связывания 3Н-ДАДЛЭ в стриатуме крыс в результате стресса. В то же время Кд этого лиганда с ОР обонятельных луковиц у НА крыс значительно возросла и стала почти в 2 раза выше, чем у ВА крыс (табл. 4). Эта разница примерно соответствует литературным данным о соотношении аффинности ДАДЛЭ к μ- и δ-ОР (Leslie, 1987, Minami, Satoh 1995, George et al., 2000). То есть можно предположить, что в обонятельных луковицах ВА крыс доминируют δ-ОР, а у НА животных в этом отделе мозга в результате стресса произошла десенситизация этих рецепторов и доминируют μ-ОР. Обонятельные луковицы играют существенную роль как в исследовательской активности грызунов, так и в формировании тревоги и депрессии, что подтверждено в экспериментах с бульбектомией (Song, Leonard, 2005). Исследовательская активность является, одним из главных факторов, определяющих поведение животных в тесте ПКЛ. Десенситизация δ-ОР в обонятельных луковицах НА, но не ВА крыс (табл. 4), по-видимому, является одной из причин того, что даларгин, более селективный лиганд δ-ОР, чем μ-ОР, оказывает анксиолитическое действие только на ВА животных (табл. 1).
Известно также, ДАДЛЭ обладает более высокой аффинностью к δ1‑ОР, чем к δ2‑ОР (Minami, Satoh, 1995). Поэтому низкую аффинность ОР обонятельных луковиц НА крыс можно объяснить доминированием там δ-ОР второго типа. Фармакологические данные, полученные в данном исследовании подтверждают это предположение: анксиолитический эффект даларгина при центральном введении у ВА крыс блокируется антагонистами обоих подтипов δ-ОР, а у НА крыс – налтрибен, селективный антагонист δ2‑ОР, не блокирует анксиолитическое действие даларгина (табл. 2). Из литературных данных следует, что роль δ2‑ОР в противотревожном действии эндогенные опиоидов более существенна, чем δ1‑ОР, поскольку налтрибен, но не BNTX оказывает анксиогенное действие на животных (Saitoh et al., 2005). Схожесть некоторых функций δ2- и μ-ОР может быть следствием того, что свойствами δ2‑ОР обладает гетеродимер μ- и δ-ОР, связанных между собой внутриклеточными С-концевыми участками (Fan et al., 2005). В зависимости от активности синтеза отдельных рецепторных белков в различных органах и, возможно, отделах мозга варьируют формы олигомеризации ОР (George et al., 2000), что также может являться причиной вариабельности биологических эффект опиоидов, в частности, их участия в регуляции тревожности.
Как упоминалось ранее, в тесте Вогеля анксиолитическое действие даларгина более выражено у НА крыс. В этом тесте оценивается способность животных преодолевать конфликт между питьевой мотивацией и болевым раздражением, сопровождающим лизание поилки. Поэтому поведение животных в тесте Вогеля в значительной степени зависит от их болевой чувствительности, в регуляции которой μ-ОР играют более существенную роль, чем δ-ОР (Gaveriaux-Ruff, Kieffer, 2002). Можно предположить, что μ-ОР доминируют не только в обонятельных луковицах (табл. 4), но и в тех отделах мозга НА крыс, которые отвечают за ноцицепцию. Если это предположение верно, то анксиолитический эффект даларгина на НА крыс в тесте Вогеля можно отчасти объяснить его обезболивающим действием, опосредуемым μ-ОР.
Таким образом, в зависимости от состояния ОР, их соотношения в различных отделах мозга, анксиолитическое действие препаратов, взаимодействующих с этими рецепторами, может по-разному проявляться в разных тестах. Как исходный уровень тревожности, так и возможность воздействия на него экзогенно вводимыми опиоидами зависят от состояния ОР и от содержания эндогенных опиоидов в их микроокружении.
К эндогенным опиоидам, в первую очередь относят опиоидные пептиды, среди которых наиболее распространены и изучены энкефалины. Энкефалины – одни из самых короткоживущих регуляторных пептидов. Период их полураспада исчисляется минутами. Поэтому активность ферментов деградации энкефалинов может играть существенную роль как в поведенческих проявлениях тревожности у животных, так и в предрасположенности и формировании некоторых форм тревожных расстройств у людей. Попытка обнаружить взаимосвязь между активностью ЭДФ в целом мозгу мышей и выраженностью поведенческих проявлений тревожности в нашем исследовании не дала убедительных результатов. Это не удивительно, поскольку существует целый ряд ферментов, участвующих в деградации энкефалинов, и их активность существенно варьирует в различных отделах мозга (Gorenstein, Snyder, 1980). Более подробное исследование активности конкретных ферментов в различных отделах мозга, ответственных за реализацию отдельных поведенческих проявлений тревожности, безусловно, дадут положительные результаты.
Удалось также обнаружить взаимосвязь между поведенческими характеристиками и активностью ЭДФ, а также содержании эндогенных опиоидов в плазме крови животных. Так, разделение крыс на 3 группы в соответствии с исходной двигательной активностью в ATS продемонстрировало, что в условиях стресса тестирования в «челночной камере» у крыс с высокой двигательной активностью (ВА) содержание лигандов ОР μ-типа в плазме крови достоверно выше, чем у остальных животных. У низкоактивных (НА) крыс обнаружено пониженное содержание лигандов ОР δ-типа (табл. 5).
Таблица 5. Активность лигандов ОР (пмоль-экв. ДАГО или ДАДЛЭ/мл) в плазме крови крыс с различной двигательной активностью в ATS
| Тип лигандов ОР | Двигательная активность животных | ||
| низкая (n=28) | средняя (n=28) | высокая (n=28) | |
| μ – (3Н-ДАГО) | 55,5±3,1 ** | 57,8±3,0 | 70,7±4,1 |
| δ – (3Н-ДАДЛЭ) | 57.6±2.4 ** ++ | 76.6±6.1 | 82.0±5.1 |
** – p<0.01 – отличие от высокоактивных крыс,
++-p<0.01, +-p<0.05 – отличие от крыс со средней двигательной активностью,
Приведены средние значения ± ошибка среднего.
Обнаружены различия в активности ЭДФ плазмы крови мышей Balb/c и C57black/6 в условиях тестирования их в ОП, эмоционально-стрессовая реакция на которое у мышей этих линий кардинально отличается. Скорость деградации лей-энкефалина (150 нМ) в плазме крови мышей линии Balb/с, проявляющих повышенный уровень тревожности в ОП, составляет 50±6 нМ/мин, что на 30% выше (p<0.05), чем у низкотревожных в этом тесте мышей C57Black/6 (38±5 нМ/мин). Можно предположить, что повышенная скорость гидролиза эндогенных опиоидных пептидов является одной из биологических причин высокой тревожности животных, а фармакологические воздействия, направленные на замедление гидролиза энкефалинов, могут оказывать анксиолитическое действие.