Скорость проникновения в клетку таких веществ, как глюкоза, глицерин, аминокислоты, не имеет линейной зависимости от разности концентраций. При определенных значениях концентрации скорость проникновения намного больше, чем следует ожидать для простой диффузии. При диффузии с подвижным переносчиком скорость переноса вещества возрастает, если молекулы (А) этого вещества образуют комплекс с молекулами (Х) вспомогательного вещества. Вспомогательное вещество обладает повышенной растворимостью в липидах. На поверхности мембраны молекулы (А) соединяются с молекулами (Х) и в виде комплекса (АХ) проходят в клетку. В клетке комплекс разрушается, молекулы вещества (А) освобождаются, а переносчик (Х) захватывает у наружной поверхности мембраны новую молекулу переносимого вещества. Процесс переноса проходит до тех пор, пока концентрация переносимого вещества не выровняется по обе стороны мембраны (рис. 3, 1).
Облегченная диффузия с фиксированным переносчиком (каналообразующим ионофором).
Цепочка молекул переносчика выстраивается внутри канала в мембране или выстилает канал. Молекула переносимого вещества (А) передвигается внутри канала от одного переносчика к другому. При этом предполагается, что пространство в канале недостаточно велико для прохождения частиц вещества, поэтому они связываются с молекулами переносчиков, передвигаясь от одного к другому (рис. 3, 2).
Ионофоры – небольшие гидрофобные молекулы, которые растворяются в липидном слое мембран и увеличивают проницаемость мембран для ионов. Большинство ионофоров синтезируется бактериями (в качестве оружия против своих конкурентов), некоторые являются антибиотиками. Широко используются в клеточной биологии.
Ионофорыделятся на следующие виды:
1.Подвижные переносчики. Имеют кольцеобразную структуру, и контактируют с липидным слоем: наружная часть – гидрофобная; внутренняя часть – гидрофильная содержит карбоксильные атомы кислорода.
2.Каналообразующие. Состоят из линейных полипептидов, которые имеют гидрофобные боковые цепи. Две таких молекулы образуют канал.
Ионные каналы (унипорт) классифицируют:
А).По типу ионов: натриевые, калиевые, кальциевые и хлорные каналы;
Б).По способу регуляции (рис. 4):
1).Потенциал–чувствительные;
2).Хемочувствительные (рецептор–управляемые);
3).Внутриклеточными веществами (ионами, физиологически активными веществами).
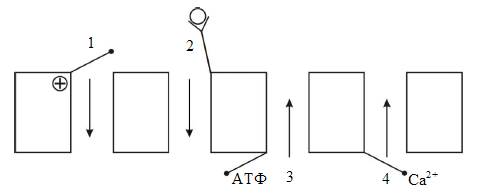
Рис. 4. Классификация ионных каналов по способу регуляции: 1 – потенциал–чувствительные; 2 – хемочувствительные (рецептор–управляемые); 3 – АТФ–чувствительные; 4 – кальций–зависимые.
Натриевые и калиевые каналы.
Размеры ионных каналов меньше одного нанометра (0,2–0,4 нм). Эти структуры обеспечивают важнейшие электрофизиологические свойства клеток и тканей, обеспечивая прохождение в 1 секунду 106–109 ионов, регулируются мембранным потенциалом и биологически активными веществами. Через ионные каналы Na+ и K+ следуют совместно с одной молекулой воды – гидратной оболочкой, хотя в растворе у иона таких оболочек может быть несколько (до 10). Таким образом, при прохождении через ионный канал происходит дегидратация катиона, на что требуется энергия. Роль этой энергии выполняет градиент ионов, так как ионный транспорт относится к пассивному виду переноса веществ через мембрану. В процессе переноса катионов должны выполняться два основных условия (фактора):
1.Стерический – совпадение размеров катиона и гидратной оболочки с размерами канала.
2.Энергетический – взаимодействие катиона с карбоксильными (отрицательно заряженными группировками самого канала).
Кальциевые каналы.
Кальциевые каналы делятся на потенциал-зависимые и рецептор-управляемые. В нормальных условиях внутри клетки [Ca2+]in = 10–7 – 10–6 M.
Через кальциевый канал проникают только двухвалентные катионы в соответствии с рядом:
pKBa2+ > pKSr2+ > pKCa2+ > pKCo2+ > pKNi2+ > pKCd2+.
Селективный фильтр кальциевого канала содержит карбоксильную группу. Все двухвалентные катионы, которые связываются с этой группой слабее, чем кальций, проникают лучше (Ba2+ и Sr2+). Катионы, которые связываются с этой группой сильнее, чем кальций, являются блокаторами кальциевых каналов (Co2+, Ni2+, Cd2+).
Регуляторный центр кальциевого канала располагается у внешнего устья. В бескальциевом растворе (с добавлением хелаторов Са2+: ЭДТА и др.) происходит модификация кальциевых каналов: они сохраняют потенциал– зависимость, но изменяют свою селективность и становятся селективными для натрия.
Процесс инактивации кальциевых каналов более сложный, чем натриевых каналов. У натриевых каналов происходит инактивация вследствие деполяризации мембраны. У кальциевых каналов инактивация зависит от силы кальциевого тока (ICa). Чем выше ICa, тем быстрее инактивация, которая развивается в результате увеличения внутриклеточной концентрации ионов кальция [Ca2+]in до 10–6 М.
Проводимость кальциевых каналов подвержена также метаболической регуляции – она зависит от содержания в цитоплазме циклического АМФ.
Кальциевые каналы подразделяют на группы:
– медленные (порог активации – 30 мВ; двухфазная инактивация; блокирование производными 1,4 – дигидропиридина);
– быстрые (порог активации – 70–60 мВ; быстрая инактивация; отсутствие чувствительности к 1,4 – дигидропиридинам).
Отличия облегченной диффузии от простой состоят в следующем:
1) перенос вещества с участием переносчика проходит значительно быстрее;
2) облегченная диффузия обладает свойством насыщения (рис. 5.8), то есть при увеличении концентрации вещества с одной стороны мембраны плотность потока вещества возрастает лишь до некоторого предела, когда все молекулы переносчика уже заняты;
3) при облегченной диффузии наблюдается конкуренция переносимых веществ в тех случаях, когда переносчик несет разные вещества; при этом одни вещества переносятся лучше, чем другие, и добавление одних веществ затрудняет транспорт других; например, из сахаров глюкоза переносится лучше, чем фруктоза, фруктоза лучше, чем ксилоза, ксилоза лучше, чем арабиноза;
4) есть вещества, блокирующие облегченную диффузию – они образуют прочный комплекс с молекулами переносчика, например, флоридзин подавляет транспорт сахаров через мембрану.

Рис. 5. Зависимость плотности потока jm вещества через мембрану в клетку от концентрации вещества снаружи клетки (СНАР) при простой (1) и облегченной (2) диффузии.
Электродиффузия
Мембранные потенциалы Между различными участками живой клетки и окружающей средой могут существовать разности потенциалов. Некоторые из этих потенциалов схематически даны на рис. 6.
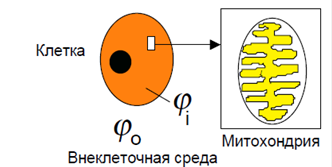
Рис. 6. Некоторые электрические потенциалы внутри живой клетки ϕo-потенциал вне клетки; ϕ i - потенциал внутри клетки; ϕ x- потенциал внутри матриксамитохондрий
Между водными фазами, разделяемыми мембранами, имеются разности потенциалов, называемые трансмембранными или же просто мембранными потенциалами. Клеточный мембранный потенциал определяют как разность потенциалов между внутриклеточным содержимым ϕ i и окружающей средой ϕo. Митохондриальный потенциал. это разность потенциалов между матриксом митохондрий ϕ x и внутриклеточной средой ϕ i. Таким образом,

Кроме трансмембранной разности потенциалов может существовать разность
электрических потенциалов между липидной фазой мембраны и омывающим водным раствором - так называемый межфазный потенциал. Если на поверхности мембраны имеются заряженные химические группы, например, остатки фосфорной кислоты, то возникает разность потенциалов между поверхностью мембраны и окружающей средой, так называемый поверхностный потенциал. Более подробно межфазные и поверхностные потенциалы будут рассмотрены позже, а сейчас мы рассмотрим, как повлияет на перенос ионов наличие на мембране трансмембранного потенциала.
На рис. 7 дан схематически профиль электрического потенциала в однородной липидной мембране, окружённой раствором солей. Из-за высокой электропроводности солевых растворов вся разность потенциалов в системе падает на липидном слое мембраны, который представляет
собой хороший электрический изолятор. Внутри мембраны, если она однородна по своим свойствам, электрический потенциал падает линейно, как это изображено на рис. 2. Это означает, что величина постоянна внутри мембраны. (Более подробно этот вопрос мы разберём в следующем разделе). По такому же линейному закону изменяется энергия иона, по мере его
продвижения поперёк мембраны.
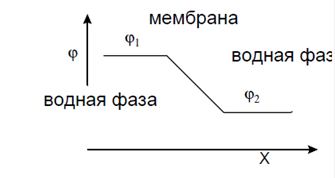
Рис.7. Профиль потенциала в однородной мембране
Эти рассуждения основаны, впрочем, на подходе к мембране, как к совершенно однородному телу. Однако липидная часть мембраны состоит всего-то из двух слоёв молекул фосфолипидов, причём размеры подвижных звеньев цепей жирных кислот в этих молекулах соизмеримы с размерами ионов, которые передвигаются внутри мембраны. Это заставляет при
рассмотрении переноса ионов в мембране отказаться от полностью макроскопического подхода к явлениям и рассматривать процессы на микроскопическом, т.е. на молекулярном уровне.