План:
1. Гуморальная регуляция
2. Гипоталамо-гипофизарная система как основной механизм нервно-гуморальной регуляции секреции гормонов.
3. Гормоны гипофиза
4. Гормоны щитовидной железы
5. Гормоны паращитовидных желез
6. Гормоны поджелудочной железы
7. Роль гормонов в адаптации организма при действии стрессовых факторов
Гуморальная регуляция - это разновидность биологической регуляции при которой информация передается с помощью биологически активных веществ, которые разносятся по организму кровью, лимфой, межклеточной жидкостью.
Гуморальная регуляция отличается от нервной:
носитель информации - химическое вещество (при нервной - нервный импульс, ПД);
передача информации осуществляется током крови, лимфы, путем диффузии (при нервной - нервными волокнами);
гуморальный сигнал распространяется медленнее (с током крови в капиллярах - 0,05 мм/с) чем нервный (до 120-130 м/с);
гуморальный сигнал не имеет такого точного «адресата» (нервный - очень конкретный и точный), воздействия на те органы, которые имеют к гормону рецепторы.
Факторы гуморальной регуляции:
 |  |  | |||
Истинные гормоны Тканевые гормоны Метаболиты
 |  | ||
«классические» гормоны
Гормоны АПУД системы
Классические, собственно гормоны - это вещества синтезируемые железами внутренней секреции. Это гормоны гипофиза, гипоталамуса, эпифиза, надпочечников; поджелудочной, щитовидной, паращитовидной, вилочковой, половых желез, плаценты (Рис. I).
Кроме эндокринных желез, в различных орынач и тканях есть специализированные клетки, которые сини шруют вещества, действующие на клетки-мишени с помощью диффузии, т. е. поступая в сровь, местно. Это гормоны паракринного действия.
К ним принадлежат нейроны гипоталамуса, которые вырабатывают некоторые гормоны и нейропептиды, а также клетки АРUD-системы, или системы захвата предшественников аминов и их декарбоксилирования. Примером могут служить: либерины, статины, нейропептиды гипоталамуса; интерстинальные гормоны, компоненты ренин-ангиотензиновой системы.
2) Тканевые гормоны секретируются неспециализированными клетками разного вида: простагландины, энкефалины, компоненты калликреин- ининовой системы, гистамин, серотонин.
3) Метаболические факторы - это неспецифические продукты, которые образуются во всех клетках организма: молочная, пировиноградная ислоты, СО2, аденозин и др, а также продукты распада при напряженном метаболизме: повышенное содержание К+, Са2+, Na+ и т.д.
Функциональное значение гормонов:
1) обеспечение роста, физического, полового, интеллектуального развития;
2) участие в адаптации организма в различных изменяющихся условиях внешней и внутренней среды;
3) поддержание гомеостаза..
Рис. 1 Железы внутренней секреции и их гормоны

Свойства гормонов:
1) специфичность действия;
2) дистантный характер действия;
3) высокая биологическая активность.
1. Специфичность действия обеспечивается тем, что гормоны взаимодействуют со специфическими рецепторами, находящимися в определенных органах-мишенях. В результате каждый гормон действует лишь на конкретные физиологические системы или органы.
2. Дистантность заключается в том, что органы-мишени, на которые действуют гормоны, как правило, расположены далеко от места их образования в эндокринных железах. В отличие от «классических» гормонов, тканевые действуют паракринно, т е. местно, недалеко от места их образования.
Гормоны действуют в очень небольших количествах, в чем и проявляется их высокая биологическая активность. Так, суточная потребность для взрослого составляет: тиреоидных гормонов - 0,3 мг, инсулина - 1,5мг, андрогенов - 5мг, естрогенов - 0,25мг и т.д.
Механизм действия гормонов зависит от их структуры
 |  | ||
Гормоны белковой структуры Гормоны стероидной структуры
Рис. 2 Механизм гормонального контроля
Гормоны белковой структуры (Рис.2) взаимодействуют с рецепторами плазматической мембраны клетки, которые являются гликопротеидами, причем специфичность рецептора обусловлена углеводным компонентом. Результатом взаимодействия является активация протеинфосфокиназ, которые обеспечивают
фосфорилирование белков-регуляторов, перенос фосфатных групп от АТФ к гидроксильным группам серина, треонина, тирозина, белка. Конечный эффект действия этих гормонов может быть - сокращение, усиление ферментных процессов, например, гликогенолиза, повышение синтеза белка, повышение секреции и т.д.
Сигнал от рецептора, с которым провзаимодействовал белковый гормон, к протеинкиназе передается с участием специфического посредника или вторичного мессенджера. Такими мессенджерами могут быть (Рис.З):
1) цАМФ;
2) ионы Са2+;
3) диацилглицерин и инозитолтрифосфат;
4) другие факторы.
Рис.З. Механизм мембранной рецепции проведения гормонального сигнала в клетке при участии вторичных посредников.

|
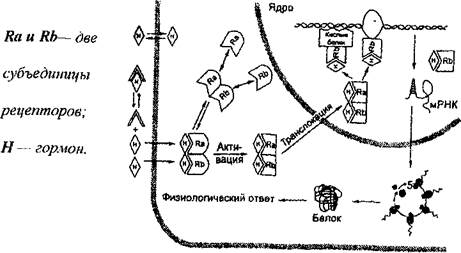
Гормоны стероидной структуры (Рис.2) легко проникают внутрь клетки через плазматическую мембрану в силу своей липофильности и взаимодействуют в цитозоле со специфическими рецепторами, образуя комплекс «гормон-рецептор», который движется в ядро. В ядре комплекс распадается и гормоны взаимодействуют с ядерным хроматином. В результате этого происходит взаимодействие с ДНК, а затем - индукция матричной РНК. Вследствие активации транскрипции и трансляции спустя 2-3 часа, после воздействия стероида наблюдается усиленный синтез индуцированных белков. В одной клетке стероид влияет на синтез не более 5-7 белков. Известно также, что в одной и той же клетке стероидный гормон может вызывать индукцию синтеза одного белка и репрессию синтеза другого белка (Рис. 4).
Рис.4 Механизм цитоплазматического (ядерного) действия стероидных гормонов.
|
Действие тиреоидных гормонов осуществляется через, рецепторы цитоплазмы и ядра, в результате чего индуцируется синтез 10-12 белков.
Рефляция секреции гормонов осуществляется такими механизмами:
1) прямое влияние концентраций субстратов крови на клетки железы;
2) нервная регуляция;
3) гуморальная регуляция;
4) нейрогуморальная регуляция (гипоталамо-гипофизарная система).
В регуляции деятельности эндокринной системы важную роль играет принцип саморегуляции, который осуществляется по типу обратных связей. Различают положительную (например, повышение сахара в крови приводит к повышению секреции инсулина) и отрицательную обратную связь (при повышении в крови уровня тиреоидных гормонов уменьшается продукция тиреотропного гормона и тиреолиберина, которые обеспечивают выброс тиреоидных гормонов).
Итак, прямое влияние концентраций субстратов крови на клетки железы идет по принципу обратных связей. Если в крови изменяется уровень вещества, который контролируется конкретным гормоном, то «слеза отвечает повышением или снижением секреции данного гормона.
Нервная регуляция осуществляется благодаря прямому влиянию симпатических и парасимпатических нервов на синтез и секрецию гормонов нейрогипофиз, мозговой слой надпочечников), а также опосредованно, «меняя интенсивность кровоснабжения железы. Эмоциональные, юихические воздействия через структуры лимбической системы, через ипоталамус - способны существенно влиять на продукцию гормонов.
Гормональная регуляция осуществляется также по принципу обратной связи: если в крови уровень гормона повышается, то в агвет на это снижается выброс тех гормонов, которые контролируют содержание данного гормона, что и приводит к уменьшению его концентрации в кроки.
Например, при повышении уровня кортизона в крови, снижается выброс АКТГ (гормон стимулирующий секрецию гидрокортизона) и как следствие
- снижение его уровня в крови. Другим примером гормональной регуляции может быть такой: мелатонин (гормон эпифиза) модулирует функцию надпочечников, щитовидной железы, половых желез т е. определенный гормон может влиять на содержание в крови других гормональных факторов.
-
Гипоталамо-гипофизарная система как основной механизм нервно-гуморальной регуляции секреции гормонов.
Функция щитовидной, половых желез, коры надпочечников регулируется гормонами передней доли гипофиза - аденогипофизом. Здесь синтезируются тропные гормоны: адренокортикотропный (АКТГ), тиреотропный (ТТГ), фолликулостимулирующий (ФС) и лютеинизирующий (ЛГ) (Рис. 5).
С некоторой условностью к тройным гормонам относится и соматотропный гормон (гормон роста), который оказывает свое влияние на рост не только прямо, но и опосредованно через гормоны - соматомедины, образующиеся в печени. Все эти тропные гормоны так названы в связи с тем, что они обеспечивают секрецию и синтез соответствующих гормонов других эндокринных желез: АКТГ -
глюкокортикоидов и минералокортикоидов: ТТГ - тиреоидных гормонов; гонадотропные - половые гормоны. Кроме того, в аденогипофизе образуется интермедии (меланоцитостимулирутощий гормон, МЦГ) и пролактин, которые обладают эффектом на периферические органы.
Рис. 5. Регуляция эндокринных желез ЦНС. ТЛ, СЛ, ПЛ, ГЛ и КЛ - оответственно, тиреолиберин, соматолиберин, пролактолиберин, гонадолиберин и кортиколиберин. СС и ПС - соматостатин и пролактостатин. ТТГ - тиреотропный гормон, СТГ - соматотропный гормон (гормон роста), Пр - пролактин, ФСГ - фолликулостимулирующий гормон, ЛГ - лютеинизирующий гормон, АКТГ - адренокортикотропный гормон

 | |||||||||||||
 |  |  |  |  | |||||||||
 | |||||||||||||
Тироксин Трийодтиронин Андрогенны Глюкортикоиды
Эстрогены
В свою очередь, высвобождение всех 7 указанных гормонов аденогипофиза зависит от гормональной активности нейронов гипофизотропной зоны гипоталамуса - в основном паравентрикулярным ядром (ПВЯ). Здесь образуются гормоны, оказывающие стимулирующее или тормозящее влияние на секрецию гормонов аденогипофиза. Стимуляторы называются рилизинг-гормонами (либеринами), ингибиторы - статинами. Выделены тиреолиберин, гонадолиберин. соматостатин, соматолиберин, пролактостатин, пролактолиберин, меланостатин, меланолиберин, кортиколиберин.
Рилизинг-гормоны освобождаются из отростков нервных клеток паравентрикулярного ядра, поступают в портальную венозную систему гипоталамо-гипофиза и с кровью доставляются к аденогипофизу.
Регуляция гормональной активности большинства желез внутренней секреции осуществляется по принципу отрицательной обратной связи: сам гормон, его количество в крови регулирует свое образование. Указанное воздействие опосредуется через образование соответствующих рилизинг- гормонов(Рис. 6,7)
 В гипоталамусе (супраоптическое ядро), кроме рилизинг-гормонов, синтезируются вазопрессин (антидиуретический гормон, АДГ) и окситоцин. Которые в виде гранул транспортируются по нервным отросткам в нейрогипофиз. Выделение нейроэндокринными клетками гормонов в кровоток обусловлено рефлекторной нервной стимуляцией.
В гипоталамусе (супраоптическое ядро), кроме рилизинг-гормонов, синтезируются вазопрессин (антидиуретический гормон, АДГ) и окситоцин. Которые в виде гранул транспортируются по нервным отросткам в нейрогипофиз. Выделение нейроэндокринными клетками гормонов в кровоток обусловлено рефлекторной нервной стимуляцией.
Рис. 7 Прямые и обратные связи в нейроэндокринно системе.
1 - медленно развивающееся и продолжительное ингибирование секреции гормонов и нейромедиаторов, а также изменение поведения и формирование памяти;
2 — быстро развивающееся, но продолжительное ингибирование;
3 - кратковременное ингибирование
Гормоны гипофиза
В задней доле гипофиза - нейрогипофизе - находятся окситоцин и вазопрессин (АДГ). АДГ влияет на клетки трех типов:
1) клетки почечных канальцев;
2) гладкомышечные клетки кровеносных сосудов;
3) клетки печени.
В почках он способствует реабсорбции воды, а значит сохранению ее в организме, снижению диуреза (отсюда название антидиуретический), в кровеносных сосудах вызывает сокращение гладких мышц, суживая их радиус, и как следствие - повышает артериальное давление (отсюда название «вазопрессин»), в печени - стимулирует глюконеогенез и гликогенолиз. Кроме этого, вазопрессин обладает антиноцицептивным эффектом. АДГ предназначен для регуляции осмотического давления крови. Его секреция увеличивается под влиянием таких факторов: повышение осмолярности крови, гипокалиемии, гипокальциемии, повышении уменьшении ОЦК, снижении артериального давления, повышении температуры тела, активации симпатической системы.
При недостаточности выделения АДГ развивается несахарный диабет: объем выделенной мочи за сутки может достигать 20л.
Окситоцин у женщин играет роль регулятора маточной активности и участвует в процессах лактации как активатор миоэпителиальных клеток. Повышение продукции окситоцина происходит во время раскрытия шейки матки в конце беременности, обеспечивая ее сокращение в родах, а также во время кормления ребенка, обеспечивая секрецию молока.
В передней доле гипофиза, или аденогипофизе, вырабатываются тиреотропный гормон (ТТГ), соматотропный гормон (СТГ) или гормон роста, гонадотропные гормоны, адренокортикотропный гормон (АКТГ), пролактин, а в средней доле - меланоцитостимулирующий гормон (МСГ) или интермедии.
Гормон роста стимулирует синтез белка в костях, хрящах, мышцах и печени. В неполовозрелом организме обеспечивает рост в длину за счет повышения пролиферативной и синтетической активности хрящевых клеток особенно в зоне роста длинных трубчатых костей, одновременно стимулируя у них рост сердца, легких, печени, почек и др органов. У взрослых он контролирует рост органов и тканей. СТГ снижает эффекты инсулина. Выброс его в кровь увеличивается во время глубокого сна, после мышечных нагрузок, при гипогликемии.
Ростовой эффект гормона роста опосредуется воздействием гормона на печень, где образуются соматомедины (А,В,С) или ростовые факторы, обуславливающие активацию синтеза белка в клетках. Особенно велико значение СТГ в период роста (препубертатный, пубертатный периоды).
В этот период агонистами ГР являются половые гормоны, увеличение секреции которых способствует резкому ускорению роста костей. Однако, длительное образование больших количеств половых гормонов приводит к противоположному эффекту - к прекращению роста. Недостаточное количество ГР приводит к карликовости (нанизм), а чрезмерное - к гигантизму. Рост некоторых костей взрослого человека может возобновиться в случае чрезмерной секреции СТГ. Тогда возобновляется пролиферация клеток ростковых зон. Что приводит к разрастанию
Кроме того, глюкокортикоиды угнетают все компоненты воспалительной реакции - уменьшают проницаемость капилляров, тормозят экссудацию, снижают интенсивность фагоцитоза.
Глюкокортикоиды резко снижают продукцию лимфоцитов, уменьшают активность Т-киллеров, интенсивность иммунологического надзора, гиперчувствительность и сенсибилизацию организма. Все это позволяет рассматривать глюкокортикоиды как активные иммунодепрессанты. Это свойство используется в клинике для купирования аутоиммунных процессов, для снижения иммунной защиты организма хозяина.
Глюкокортикоиды повышают чувствительность к катехоламинам, повышают секрецию соляной кислоты и пепсина. Избыток этих гормонов вызывает деминерализацию костей, остеопороз, потерю Са2+ с мочой, снижают всасывание Са2+. Глюкокортикоиды влияют на функцию ВНД - повышают активность обработки информации, улучшают восприятия внешних сигналов.
Минералокортикоиды (альдосгерон, дезоксикортикостерон) участвуют в регуляции минерального обмена. Механизм действия альдостерона связан с активацией синтеза белка, участвующего в реабсорбции Na+ - Na+, Кч-АТФазы. Повышая реабсорбцию и снижая ее для К+ в дистальных канальцах почки, слюнных и половых железах, альдостерон способствует задержке №' и СГ в организме и выведению К+ и Н из организма. Таким образом, альдостерон является натрийсберегающим, а также калийуретическим гормоном. За счет задержки Иа\ а вслед за ним и воды, он способствует повышению ОЦК и, как следствие, повышению артериального давления. В отличие от глкжокортикоидов, минералокортикоиды способствуют развитию воспаления, т.к. повышают проницаемость капилляров.
Половые гормоны надпочечников выполняют функцию развития половых органов и появление вторичных половых признаков в тот период, когда половые железы еще не развиты, т е. в детском возрастем также в пожилом возрасте.
Гормоны мозгового слоя надпочечников - адреналин (80%) и норадреналин (20%) - вызывают эффекты во многом идентичные активации нервной системы. Их действие реализуется за счет взаимодействия с а- и (3- адренорецепторами. Следовательно, им присуща активация деятельности сердца, сужение сосудов кожи, расширение бронхов и т.д. Адреналин влияет на углеводный и жировой обмен, усиливая гликогенолиз и липолиз.
Катехоламины участвуют в активации термогенеза, в регуляции секреции многих гормонов - усиливают выброс глюкагона, ренина, гастрина, паратгормона, кальцитонина, тиреоидных гормонов; снижают выброс инсулина. Под влиянием этих гормонов повышается работоспособность скелетных мышц, возбудимость рецепторов.
При гиперфункции коры надпочечников у больных заметно изменяются вторичные половые признаки (например, у женщин могут появляться мужские половые признаки - борода, усы, тембр голоса). Наблюдаются ожирение (особенно в.области шей, лица, туловища), гипергликемия, задержка воды и натрия в организме и др.
Гипофункция коры надпочечников вызывает болезнь Аддисона – бронзовый оттенок кожи (особенно лица, шеи, рук), потеря аппетита, рвота, повышенная чувствительность к холоду и боли, высокая восприимчивость к инфекциям, повышенный диурез (до 10 л мочи за сутки), жажда, снижение работоспособности.