Под влиянием внешних факторов частота аллелей постоянно меняется, и без этого невозможно элементарное эволюционное явление. Какие факторы оказывают влияние на популяцию и в чем конкретно проявляется их действие?
Элементарные эволюционные факторы. Прежде всего для эволюции необходимо наличие факторов, поставляющих в популяции эволюционный материал. Эту роль выполняют мутационный процесс, комбинативная изменчивость, поток, генов, периодические случайные колебания численности популяций. Имея различную природу, они действуют случайно и ненаправленно, поставляя на эволюционную арену разнообразные генотипы.
Важное значение для эволюции имеют факторы, создающие барьеры, препятствующих скрещиванию,— это различные формы изоляции, закрепляющие любые различия в наборах генотипов в разных частях популяции.
Наконец, необходимо наличие естественного отбора — фактора, направляющего эволюционный процесс.
Все эти факторы оказывают давление на популяцию, приводят к возникновению элементарного эволюционного явления.
I. НЕНАПРАВЛЕННЫЕ ФАКТОРЫЭВОЛЮЦИИ.
1. Давление мутационного процесса. Мы знаем, что мутации являются элементарным эволюционным материалом. М утационный процесс оказывает ощутимое давление на генофонд популяции
Частоты возникновения отдельных новых мутаций обычно относительно низки: одна мутация на 10000—1000000 особей (гамет) в поколении. Но в связи с большим числом генов (их десятки тысяч у высших форм) общая частота всех возникающих мутаций у живых организмов достаточно высока: от нескольких единиц до нескольких десятков процентов. У дрозофилы до 25% гамет на одно поколение несут мутации..
Мутантная особь – альбинос

Мутации по окраске шерсти у лисицы обыкновенной (чёрно-бурая и платиновая)



Мутации и их комбинации в эволюции. Значительная часть вновь возникающих мутаций снижает жизнеспособность особей по сравнению с исходной нормой. Большинство отклонений от нормы должны оказаться «вредными» уже потому, что случайное изменение сложного механизма в подавляющем большинстве случаев не может его улучшить. Однако при переходе в гетерозиготное состояние многие мутации не только не снижают жизнеспособность несущих их особей, но и в некоторых случаях даже повышают ее. Эта особенность мутаций хорошо известна в практике животноводства и растениеводства: часто две мутантные формы, мало жизнеспособные порознь, объединенные в потомстве, обладают ценными хозяйственными свойствами.
Как показали опыты Н.В. Тимофеева-Ресовского, жизнеспособность одних и тех же мутаций зависит от того, в комбинации с какими аллелями других генов они оказываются. В зависимости от генного окружения («генотипической среды») жизнеспособность одной и той же мутации может изменяться.
Небольшой процент мутаций с самого начала может принести в конкретных условиях не вред, а пользу для особи. Как бы ни была мала доля таких мутаций, они, в грандиозных временных масштабах процесса эволюции, сами по себе могут сыграть заметную положительную роль.
Мутационный процесс носит случайный и ненаправленный характер. Возникая, разнообразные мутации изменяют исходные признаки в различных направлениях, осуществляя в классической форме дарвиновскую неопределенную изменчивость.
Ненаправленность мутационного процесса отчетливо проявляется при искусственном мутагенезе, когда внешние воздействия, вызвавшие мутацию, точно известны. Так, хорошо известно мутагенное действие проникающей радиации. Возникающие под ее влиянием мутации могут затрагивать самые различные особенности организма (цвет глаз, степень развития и особенности строения крыльев у мух), вызывать самые разные наследственные заболевания у человека и других организмов. Но эти изменения в подавляющем большинстве никак не связаны с защитой организма от радиации, т. е. не имеют приспособительного значения.
Таким образом, мутационный процесс лишь поставляет материал для новых и новых эволюционных изменений, т. е. играет важнейшую роль «поставщики» элементарного эволюционного материала. Сам по себе, без участия других факторов эволюции, в первую очередь естественного отбора, мутационный процесс не может привести к направленному изменению генофонда популяции.
2. Комбинативная изменчивость. Возникнув, отдельные мутации оказываются в соседстве с другими мутациями, входят в состав новых генотипов, т. е. возникает множество сочетаний аллелей. Расчеты показывают, что любая особь генетически уникальна. Так, если допустить, что в каждой паре гомологичных хромосом имеется только одна пара аллельных генов, то для человека, у которого гаплоидный набор хромосом равен 23, число возможных генотипов составит 3(в степени 23). Такое огромное количество генотипов в 20 раз превышает численность всех людей на Земле. Однако в действительности всё ещё сложнее. Поэтому количество возможных генотипов выражается астрономическим числом, и можно с уверенностью утверждать, что возникновение двух одинаковых людей практически невероятно. Таким образом, обмен генами вследствие перекреста хромосом в мейозе и случайность слияния гамет в половом процессе — два фактора, на которых основано многообразие комбинаций
3. Поток генов. Важным источником изменчивости служит поток генов — обмен генами между популяциями одного вида в результате свободного скрещивания их особей. Часть особей-мигрантов одной популяции проникает в другую, и их гены включаются в генофонд этой популяции. При скрещивании особей разных популяций генотипы потомства будут отличаться от генотипов обоих родителей. В данном случае происходит перекомбинация генов на межпопуляционном уровне.
Популяционные волны.
Нашествия полевок, мышей, саранчи известны человечеству с библейских времен. Еще Аристотель оставил описание «расцвета и падения» мышиной популяции. Он отмечал, что нашествие грызунов было бедствием, сравнимым с чумой. Они чудовищно размножились, уничтожили урожай и свою собственную «пищевую базу» и в конце концов исчезли, как сквозь землю провалились. В Древней Руси годы массового размножения грызунов называли годами «мышиной напасти», о них даже упоминается в летописях. В наши дни такого масштаба мышиной напасти уже не бывает, потому что численность грызунов люди научились регулировать. Однако колебания численности, особенно хорошо заметные у мышевидных грызунов и других видов с коротким жизненным циклом и быстрой сменой поколений, характерны для всех популяций растений и животных.
В 1905 г. С. С. Четвериков опубликовал работу под заглавием «Волны жизни», в которой раскрыл значение колебаний численности популяций — популяционных волн, или «волн жизни», для эволюции.
Нашествие саранчи

 Размножение иван-чая на пожарище
Размножение иван-чая на пожарище
Размножение кроликов в Австралии

История популяционных волн у мышей

Причины колебания численности популяций. У быстро размножающихся видов наблюдается периодическое чередование подъемов и спадов численности — популяционные циклы. Так, циклы полевок, леммингов и других мышевидных грызунов длятся обычно 4 года. За этот срок количество животных возрастает от ничтожного до максимального, затем падает почти до нуля и начинается новый цикл. Каковы причины такой периодичности? Трудно дать исчерпывающее объяснение. По-видимому, немалую роль в этом процессе играют хищники, численность которых колеблется пропорционально росту и убыли популяции грызунов. Например, чем больше полевок, тем больше птенцов выводят совы. Когда популяция мышей идет на убыль, совы снимаются с гнезд и улетают в поисках пищи в другие места. На старом месте остается ровно столько птиц, сколько может прокормиться. Пустельги, луни, канюки и другие постоянно обитающие в одном месте птицы в период рассвета мышиной популяции выкармливают всех вылупившихся птенцов, а в голодный год много птенцов гибнет. Однако хищничество -- только одна из многих причин колебания численности. Хищник съедает не более того, что ему необходимо, и бессилен справиться с полчищами животных в период их массового размножения. Резкие колебания численности грызунов могут быть связаны и со вспышками эпидемий.
Причины популяционных волн в одних случаях менее известны, в других — более изучены и объяснимы. Так, хорошо известно, что урожай еловых шишек повышается после теплого сухого лета, а это, в свою очередь, положительно сказывается на росте популяции белок.
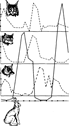 Динамика численности зайцев и хищников
Динамика численности зайцев и хищников
Резкие непериодические падения численности возникают в результате засухи, пожара, наводнения и других природных «катастроф» При этом всегда неизбежно создаются исключительно благоприятные условия для развития одних организмов, неблагоприятные - для других. Например, на месте лесных пожаров буйно разрастается иван-чай. Его численность увеличивается на протяжении нескольких лет. затем это растение постепенно вытесняется другими травами, кустарниками, деревьями.
Резкие вспышки численности видов наблюдаются при их попадании в новые подходящие для жизни условия. Достаточно вспомнить последствия расселения ондатры в Европе и в СССК, завоевание кроликами Австралии. Однако через несколько поко^-лений новый для данного биогеоценоза вид становится жертвой новых для него хищников, новых болезней, к которым не выработан иммунитет В результате вслед за небывалым подъемом численности неизбежно наступает период глубокого ее "адения_1 ак было в СССР с ондатрой в 50—60-х годах, так было в 1987-1988 гг на озере Севан с акклиматизированным здесь сигом.
Каким бы ни был механизм популяционных волн, ясно, что на численность популяции могут влиять одновременно многие факторы которые неизбежно приводят к периодическим или непериодическим, сезонным или годовым изменениям численности любого из известных видов животных и растении.
Значение «волн жизни» для эволюции. С возрастанием численности популяции увеличивается вероятность появления новых мутаций и их комбинаций. Если в среднем один мутант появляется на 100 тыс особей, то при возрастании численности популяции в 100 раз число мутантов увеличивается во столько же раз. После спада численности сохранившаяся часть особей популяции по генетическому составу будет значительно отличаться от ранее многочисленной популяции: часть мутаций совершенно случайно исчезнет вместе с гибелью несущих их особей, а некоторые мутации, также случайно, резко повысят свою концентрацию. Естественно что после восстановления численности до нормального уровня генофонд популяции будет уже иной.
Таким образом, популяционные волны сами по себе не вызывают наследственную изменчивость, а только способствуют изменению частот аллелей и генотипов. Действуя совершенно иначе, чем мутационный процесс, «волны жизни:, являются вместе с ним фактором-поставщиком эволюционного материала, выводящим ряд генотипов совершенно случайно и ненаправленно на эволюционную арену.
Генофонд популяции меняется в результате популяционных волн

Генетический дрейф.
Возрастание численности вида, как правило сопровождается расширением его ареала. Попав за пределы ареала, небольшие популяции могут закрепиться в новых местах и там попадут в совершенно новые условия. В таких маленьких популяциях влияние случайных факторов может быть очень сильно и резко изменит генофонда.
Случайное ненаправленное изменение частот аллелей в малых популяциях назвали генетическим дрейфом.

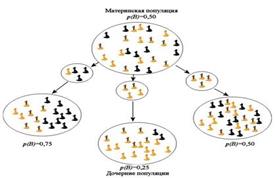
Генофонд малочисленной популяции с самого начала беднее генофонда родительского вида, и подбор аллелей в нем случаен (в результате случайного подбора особей). Поскольку естественный отбор происходит эффективно при достаточно большой численности особей, дальнейшая судьба генофонда малой популяции определяется главным образом действием различного рода случайных факторов.
Изоляция
Под изоляцией понимается возникновение любых барьеров, нарушающих свободное скрещивание. Это элементарный эволюционный фактор, действующий вначале в пределах популяции, на микроэволюционном уровне.
В зависимости от самих барьеров выделяют два основных типа изоляции: первичную и вторичную = репродуктивную.
Первичная изоляция связана с изменениями в ландшафте: образование преград в виде рек, горных хребтов, лесных массивов и т. д. В результате свободное скрещивание сухопутных особей затрудняется из-за водных барьеров, а особей, обитающих в воде, — из-за барьеров суши. Возвышенности изолируют равнинные популяции, а равнины — горные популяции.
Мыши были завезены из Европы на Фарерские острова около 200 лет назад – достаточно изолированная популяция
Первичная изоляция может возникнуть и без заметных географических барьеров, просто в силу большого расстояния между популяциями одного вида. Часто причиной является деятельность человека в биосфере. Например, еще в начале нашего века ареал соболя был сплошным и занимал значительную часть Евразии, но уже в 20—30х годах в связи с интенсивным перепромыслом этого зверька его ареал приобрел мозаичную структуру: распался на отдельные участки, разделенные значительным расстоянием, препятствовавшим свободному скрещиванию удаленных друг от друга популяций. В 40—50е годы ареал соболя в СССР был восстановлен.
Изоляция может быть абсолютной. Например, популяции песцов, обитающие на Командорских островах и на Камчатке, не обмениваются особями, так как разделены незамерзающим проливом. Однако изоляция зависит не только от пространственного расположения географических барьеров, но и от радиуса индивидуальной активности — расстояния, на которое способны перемещаться особи в период размножения. Если радиус индивидуальной активности невелик по сравнению с размерами популяции и расстоянием между соседними популяциями, то степень, или давление, изоляции будет большим. Например, популяции наземных моллюсков, удаленные друг от друга на 200—400 м, будут существовать изолированно, так как их индивидуальная подвижность ограничена.
Игуана – малоподвижное животное, радиус активности около 700 м.

А песцы могут мигрировать по льдам Арктики на сотни километров, и в результате изоляция песцов острова Врангеля от популяций песцов Чукотки очень невелика. Если радиус индивидуальной активности позволяет особям соседних популяций преодолевать расстояние между ними (степень изоляции незначительная), то эти популяции очень скоро приобретут общие свойства из-за полного перемещения генетического материала. Так, утки одного вида занимают ареал, равный чуть не всей Европе, и свободно скрещиваются между собой. Предполагают, что все они относятся к одной популяции или очень немногим популяциям.
Вторичная, или репродуктивная, изоляция. Биологическая (репродуктивная) изоляция приводит к нарушению скрещивания или препятствует воспроизведению нормального потомства.
Например скрещивание нарушается из-за того, что встреча партнеров мало вероятна – у них различны сроки спаривания, ритуал ухаживания, брачные песни и т. д.
Каждый вид светлячков имеет свою динамику вспышек свечения


У растений одного вида многочисленны случаи несовпадения времени цветения и опыления. В озере Севан существовало несколько популяций форели, отличающихся сроками и местами нереста. Может быть разным строение полового аппарата.
Также из-за разницы в хромосомных наборахпоявляютсягибриды с пониженной жизнеспособностью или гибриды не дают своего потомства.
Эволюционное значение изоляции. Нарушая свободу скрещивания, она закрепляет различия в частотах встречаемости различных генотипов (тогда как свободное скрещивание ведет к сглаживанию различий). Однако ее давление на популяцию так же ненаправленно, как и давление факторов — поставщиков эволюционного материала.
Ненаправленность названных факторов эволюционного процесса — главная черта, отличающая их от естественного отбора — направленного элементарного эволюционного фактора. Факторы-поставщики эволюционного материала изменяют частоты аллелей, а изоляция усиливает их действие независимо от того, ведет это к повышению или к понижению приспособленности организмов к условиям существования.