Общей количественной мерой различных форм движения материи является энергия. Согласно первому закону термодинамики, энергия как и материя в целом не возникает и не исчезает, а лишь переходит из одной формы существования в другую. Раскрытие сущности таких переходов позволяет человеку управлять этими явлениями и использовать на практике для решения важнейших проблем.
Примеров таких взаимопереходов достаточно: электрическая энергия ↔ электромагнитная (на этой основе изобретен электродвигатель и динамомашина);
энергия света ↔ электрическая энергия (солнечные батареи, осветительные приборы); электрическая энергия ↔ энергия тепловая (нагревательные приборы, тепловые электростанции).
Энергетические преобразования лежат в основе функционирования и биологических систем. Более того, в некоторых биологических системах имеет место исключительно важное (для всех организмов на Земле), энергетическое преобразование – трансформация энергии света в химическую энергию.
Уникальность такого преобразования энергии состоит в том, что, во-первых, химическая форма энергии может долго храниться и в случае необходимости постепенно расходоваться растениями. А во-вторых, что самое главное, эта энергия помимо того, что она может быть использована на различные физико-химические процессы растения, используется всеми живыми существами на Земле (за исключением хемосинтетиков) и для них является единственным источником. Вполне естественно, что, прежде чем стала возможной такая трансформация энергии, органическая материя должна была достигнуть определенного уровня развития. Как известно, данное превращение происходит в процессе фотосинтеза и по праву занимает в нем центральное место.
Условием существования растительного организма как открытой биологической системы служит обмен энергией, веществом и информацией, как между частями системы, так и с окружающей средой. Это позволяет ему избегать термодинамической смерти в определенный промежуток времени. Ключевую роль в этих преобразованиях играют мембранные комплексы. Особо следует отметить роль мембран хлоропластов, которые поглощают внешнюю неустойчивую энергии квантов света и преобразовывают ее во внутреннюю устойчивую энергию химических связей органических веществ. Такое энергетическое преобразование происходит через ряд этапов.
Первоначально внешняя неустойчивая энергия квантов света преобразуется во внутреннюю энергию электронного возбуждения. При этом она частично стабилизируется. Это энергетическое преобразование происходит на фотофизическом этапе фотосинтеза. В дальнейшем энергия электронного возбуждения накапливается первично в лабильных метаболически активных соединениях с высоким энергетическим потенциалом – АТР и в форме восстановленных кофакторов NADPH2 , FdH2. Эти преобразования имеют место на фотохимическом этапе фотосинтеза.
Процесс образования АТР в реакциях фотосинтеза был открыт в 1954 г. Арноном и получил название фотосинтетического фосфорилирования (ф/ф).
В исследованиях реакций ф/ф-я можно выделить два периода. Первый (1954 − 1958 гг.) был направлен на разработку высокочувствительных методов изучения светового синтеза АТР. Второй период посвящен изучению механизма этих реакций. В самом начале этого периода была выдвинута гипотеза о связи синтеза АТР с транспортом электронов, донором которых является возбужденный светом хлорофилл.
Было показано:
1. Процесс ф/ф-я подчиняется основным положением термодинамики.
2. При фотоиндуцируемом транспорте электронов по ЭТЦ, происходит ряд сопряженных о/в реакций. Энергия, которая выделяется при этом, используется на синтез АТР.
3. Были выделены компоненты ЭТЦ, определены их о/в потенциалы и место локализации.
По ЭТЦ электроны могут спонтанно перемещаться от электроотрицательных к электроположительным компонентам цепи. Этот перенос сопровождается выделением энергии. Количество выделяемой энергии прямо пропорционально разности о/в потенциалов двух систем. Расчеты показывают, что для образования макроэрга достаточна, разность о/в потенциала 0,25в (0,15в). Эта энергия при наличии сопрягающих механизмов может быть использована для синтеза АТР.
При расшифровке ЭТЦ выяснилось, что ток электронов движется от воды о/в +0,81в к NADP о/в -0,32в, т.е. против термодинамического градиента. Поэтому спонтанный ток электронов в этой системе не возможен. Для того чтобы направить 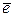 против термодинамического градиента, ему необходимо сообщить
против термодинамического градиента, ему необходимо сообщить
дополнительную энергию. Эту энергию  получает, поглотив hn.
получает, поглотив hn.
Использование различных кофакторов и ингибиторов, а также монохроматического света позволила выделить 3 типа ф/ф-я: циклическое, нециклическое и псевдоциклическое.
Нециклическое ф/ф-е является основным процессом запасания энергии, протекающее в аэробных условиях. Отличительными особенностями нециклического ф/ф-я являются:
1. Участие двух фотосистем.
2. Нециклический транспорт электронов от Н2О к NADP.
3. Образование двух энергетических эквивалентных АТР и NADPH2 которые используются в темповой фазе фотосинтеза.
4. Происходит фотоокисление Н2О и выделение О2 и образование Н2О.
Образование на свету энергетических эквивалентов АТР и NADPH2 происходит благодаря ряду о/в реакций, которые в своей совокупности и составляют фотохимический этап.
Нециклическое ф/ф-е начинается с того, что энергия квантов света, поглощенная I пигментной системой, которая стекается к длинноволновой форме хлорофилла Р700, которая является ловушкой и связана с реакционным центром. После поглощения определенной порции энергии,  молекулы Р700 переходят из S0 в S1 и далее может передаваться переносчикам ЭТЦ. При переносе
молекулы Р700 переходят из S0 в S1 и далее может передаваться переносчикам ЭТЦ. При переносе 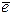 по ЭТЦ происходит ряд о/в реакций, в результате которых выделяется энергия. В местах достаточного ее выделения и при наличии сопрягающих механизмов образуется АТР.
по ЭТЦ происходит ряд о/в реакций, в результате которых выделяется энергия. В местах достаточного ее выделения и при наличии сопрягающих механизмов образуется АТР.
Первым компонентом ЭТЦ воспринимающим 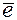 от Р700 является не идентифицированный переносчик Z о/в потенциал которого -0,55. Пигмент Р700 в основном состоянии имеет о/в +0,43 в. Следовательно, спонтанный ток невозможен. Однако после поглощения hn молекулой Р700 её о/в падает до - о,6 в и она отдает свой
от Р700 является не идентифицированный переносчик Z о/в потенциал которого -0,55. Пигмент Р700 в основном состоянии имеет о/в +0,43 в. Следовательно, спонтанный ток невозможен. Однако после поглощения hn молекулой Р700 её о/в падает до - о,6 в и она отдает свой  . О/в потенциал всех остальных переносчиков более электроположителен, что обеспечивает в дальнейшем спонтанный ток . Следующим компонентом ЭТЦ является Fd (ферредоксин) - железосодержащий белок с о/в потенциалом -0,43 в, железо в молекуле ферредоксина находится в негеминовой форме. От Fd
. О/в потенциал всех остальных переносчиков более электроположителен, что обеспечивает в дальнейшем спонтанный ток . Следующим компонентом ЭТЦ является Fd (ферредоксин) - железосодержащий белок с о/в потенциалом -0,43 в, железо в молекуле ферредоксина находится в негеминовой форме. От Fd  переносится на NADР (о/в -0,32 в). Перенос возможен благодаря ферменту ферредоксин- NADP -редуктазы, коферментом которого являются флавопротеиды.
переносится на NADР (о/в -0,32 в). Перенос возможен благодаря ферменту ферредоксин- NADP -редуктазы, коферментом которого являются флавопротеиды.
После выброса пигмент Р700 имеет вид ионизированной молекулы (её о/в потенциал вновь становится -0,43 в), и она готова принять , Образовавшуюся дырку заполняет электрон второй фотосистемы.
Работа второй ФС начинается также с поглощения hn пигментной системой. Далее энергия стекается к длинноволновой форме хлорофилла Р680. Возбужденный электрон передается не идентифицированному переносчику (Q), о/в потенциал последующих переносчиков более электроположительный, поэтому в дальнейшем перенос  происходит спонтанно. От переносчика Q передается на пластохинон (о/в +0,11 в) активной частью молекулы является хинон.
происходит спонтанно. От переносчика Q передается на пластохинон (о/в +0,11 в) активной частью молекулы является хинон.
 +2Н+ +2е-
+2Н+ +2е-

Однако, для восстановления хинона до гидрохинона необходимы не только 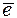 , но и Н+, которые он получает из стромы хлоропласта. От пластохинона поступает на цитохромы. Цитохромы - металлопротеиды, простетической группой которых являются железо-порфирины. Известно около 20 цитохромов, которые отличаются радикалами в порфириновом ядре.
, но и Н+, которые он получает из стромы хлоропласта. От пластохинона поступает на цитохромы. Цитохромы - металлопротеиды, простетической группой которых являются железо-порфирины. Известно около 20 цитохромов, которые отличаются радикалами в порфириновом ядре.
Первым цитохромом, воспринимающим  от пластохинона является Цит559.
от пластохинона является Цит559.
Непосредственное участие в о/в реакциях принимает железо, входящее в состав гема.

 Fe+3 +
Fe+3 +  Fe+2
Fe+2
Для восстановления железа Н+ не нужны и они выбрасываются во внутритилакоидное пространство. Гидрохинон при этом вновь окисляется до хинона. Последующим переносчиком является Цитf (о/в+0,36 в), активной частью которого также является железо. Конечным компонентом ЭТЦ ФС II является пластоцианин – медьсодержащий белок (о/в +0,637 в). От пластоцианина передается на P700 и заполняет дырку.
При переносе  по ЭТЦ происходит ряд о/в реакций с выделением энергии. В тех местах, где ее достаточно для образования макроэргических связей, образуется АТР из АDР и Рi.
по ЭТЦ происходит ряд о/в реакций с выделением энергии. В тех местах, где ее достаточно для образования макроэргических связей, образуется АТР из АDР и Рi.
Для заполнения дырки P680 поступает из воды. Поэтому работа ФС II связана с разложением (фотоокислением) Н2О и выделением О2.
Впервые выделение 02 наблюдал Хилл. Он добавлял к изолированным хлоропластам экзогенные акцепторы (эндогенные Fd и NADР+ растворимы в воде и вымываются при их выделении) ДХФИФ K4[Fe(CN)6] освещал эту систему и наблюдал, что восстановление акцепторов сопряжено с одновременным выделением О2. Именно этот процесс носит название реакции Хилла.
Большое значение в разложении Н2О играет Мп (марганцевые протеиды) и также Ca и Cl.
Q
↑
P680 ← y ← Mn ← Н2О
Н2О  H+ +
H+ +  + (·ОН)
+ (·ОН)
 заполняет дырку Р670.
заполняет дырку Р670.
2 Н+ - присоединяется к NADP и дают NADPH2
2 (∙ОН) → Н2О +  O2
O2
В настоящее время установлено, что Мn является обязательным компонентом, участвующим в фотоокислении воды. Более того, в лаборатории академика А.Е.Шилова была найдена система, состоящая из Мn и Рутения. В водном растворе под действием света она выделяла O2. А.Е.Шилов обратил внимание на то, что Мn, является единственным элементом, по крайней мере из используемых в живой природе, который способен изменять степень окисления на пять единиц, образуя в воде устойчивые частицы.
Попутно можно заметить, что в этой же лаборатории была смоделирована система, которая под действием ультрафиолетовых лучей восстанавливала Н2.
Чтобы отнять 2  нужно 4hn, но чтобы выделилась 1 молекула СО2 необходимо 8 hn- квантовый расход – это количество квантов, необходимое для восстановления 1 молекулы СО2 до углеводов.
нужно 4hn, но чтобы выделилась 1 молекула СО2 необходимо 8 hn- квантовый расход – это количество квантов, необходимое для восстановления 1 молекулы СО2 до углеводов.
Общая схема нециклического ф/ф-я:
8 hn
2ADP + 2Pi +2NADP + 2H2O → 2ATP +2NADPH +2H+ +O2
Кофакторы: K4[Fe(CN)6], NADP+ + (Fd)
Ингибиторы: моно- и диуроны, гидроксиламин.
Циклическое ф/ф-е: имеет следующие особенности:
1. Циклический транспорт .
2. Участие в процессе только ФС I.
3. Энергия запасается только в виде АТР.
4. Процесс может протекать в анаэробных условиях.
Циклическое ф/ф-е правильнее всего рассматривать как один из путей процесса образования АТР. В зависимости от внешних условий ток может переключаться по циклическому или нециклическому пути. Роль регулятора, в этом процессе выполняет Fd. (Метаболическая вилка)
Циклическое ф/ф-е начинается также как и нециклическое. Электрон от Р700 поступает к Z, а от него на Fd. Далее ток может идти двумя путями. В случае нециклического ф/ф-я он сбрасывается на NADP. При циклическом ф/ф-ии  через переносчики Цf, вновь возвращается к молекуле P700.
через переносчики Цf, вновь возвращается к молекуле P700.
При транспорте  по ЭТЦ в местах с достаточной разностью о/в между переносчиками выделяется необходимое количество энергии для образования АТР из ADP и Рi. Однако эта энергия может быть запасена в виде ~АТР, только в том случае, если имеется фермент сопряжения CF, в противном случае, она будет выделена в виде тепла.
по ЭТЦ в местах с достаточной разностью о/в между переносчиками выделяется необходимое количество энергии для образования АТР из ADP и Рi. Однако эта энергия может быть запасена в виде ~АТР, только в том случае, если имеется фермент сопряжения CF, в противном случае, она будет выделена в виде тепла.