Отмеченная ранее малая вязкость солевых растворов имеет ту оборотную сторону, что она не позволяет заранее (как это делают для растворов сахарозы) создавать в центрифужной пробирке градиент концентрации соли. Этот градиент очень быстро будет разрушен за счет диффузии ионов соли. Зато, ввиду тяжести молекулы CsCI, при длительном и неизменном по скорости вращении первоначально однородного раствора соли устанавливается динамическое равновесие между седиментацией самих молекул соли и их диффузией. В результате образуется (в ходе самого центрифугирования!) некий, нелинейный градиент концентрации CsCI, a следовательно, и плотности его раствора по высоте пробирки. Вполне пригодный для фракционирования биологических макромолекул или частиц по их плавучей плотности.
Установление градиента плотности раствора CsCI в ходе центрифугирования — процесс долгий. Он занимает, как минимум, 60—70 часов. Его можно несколько ускорить, если предварительно заполнить пробирку тремя равными порциями растворов CsCI различной плотности: средней для среднего слоя заполненной пробирки и, соответственно, увеличенной или уменьшенной на половину от величины различия плотностей, которые ожидают получить для жидкой среды центрифугирования у дна пробирки и на ее мениске.
С этой же целью равновесное ультрацентрифугирование проводят иногда в угловых роторах. Распределение плотностей раствора CsCI во время центрифугирования идет все равно в направлении радиуса вращения, но значительно быстрее, поскольку в этом направлении наклонно стоящая пробирка намного короче (см. рис. 48а). На том же рисунке условно-дискретными зонами показано расположение слоев различной плотности CsCI к концу центрифугирования (различная штриховка) и находящиеся
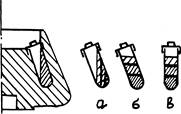
Рис. 48
между ними полосы разделившихся частиц (жирными линиями на рис. 486). Положение этих зон и полос в пробирке, извлеченной из ротора по окончании центрифугирования показано на рис. 48 в. В течение некоторого времени, пока диффузия не размоет эту картину, раздельное положение полос вещества сохраняется.
В качестве примера приведу результат очистки равновесным центрифугированием плазмид от примеси основной массы ДНК бактерии (Currier, Nester, 1976). Основную массу ДНК бактерии — хозяина удаляли денатурацией при рН12,2 и переводом ее в фенольную интерфазу в присутствии 0,5 М NaCI (см. гл. 3, § 2). Плазмидная ДНК оставалась в водной фазе, где она легко ренатурируется. Дальнейшую ее очистку вели равновесным центрифугированием в CsCI с добавлением бромистого этидия для окраски полос. Использовали 48% -ный раствор CsCI, чему соответствовала его начальная плотность 1,55 г/см3. (Столь малое значение этой плотности обусловлено уменьшением плавучей плотности ДНК обоих типов за счет связи с красителем.) Центрифугировали при температуре 14°С в угловом роторе (Spinco-40) при 35 000 об/мин в течение 60 часов. На графике рис. 49 показано надежное отделение ДНК плазмид (пик 1) от остатков основной ДНК бактерии (пик 2).

Литература
1. Музя Г.И., Куликов В.И., Ванько Л.В., Сухих Г.Т. Роль фосфолипидного фактора активации тромбоцитов в репродукции // Акушерство и гинекология -1996. -3. -С.12-16.
2. Музя Г.И., Орлов С.М., Бердышев Е.В., Куликов В.И. Взаимодействие 1-О-алкил-2-метокси-sn-глицеро-3-фосфохолина и фактора активации тромбоцитов с клетками крови и опухолевыми клетками // Биохимия.-1994.-Т. 56.-С.-1054-1061.
3. Мысляева Т.Г. Динамика электролитов в процессе обезвоживания организма. Всесоюзная конференция. - Ленинград.1978.