ВПСП,РП и ГП обеспечивают возникновение ПД в нервных элементах или мышечных клетках, что достигается за счет действия электрического поля локального потенциала, деполяризующего клеточную мембрану. Когда деполяризация достигает 50% Екр, активируются Na+-каналы, в результате чего преобладает движение ионов Na+ в клетку и развивается дальнейшая ее деполяризация. Если амплитуда ЛП достаточна для обеспечения Екр (100% Екр), то возникает ПД в соответствующих структурах клеток возбудимых тканей. Например, ПКП обеспечивает возникновение ПД в прилежащем к концевой пластинке участке мышечного волокна, ГП обеспечивает возникновение ПД в нервном волокне (в миелиновом волокне - это первый перехват Ранвье). Если амплитуда ЛП достаточна для обеспечения возникновения ПД, то ЛП сливается с ПД и является началом первой его фазы - деполяризации. Если же амплитуда ЛП недостаточна, он затухает в структурах, где возник (постсинаптические мембраны, мембраны сенсорных рецепторов). Это затухание происходит обычно вследствие преобладающего движения ионов К+ из клетки по каналам утечки ионов, которые всегда открыты, а также - по каналам, которые активировались медиатором (постсинаптические мембраны) или по каналам мембран сенсорных рецепторов, которые активировались действием соответствующего адекватного раздражителя, например, света, прикосновения. Для передачи возбуждения на большие расстояния необходимо формирование ПД.
Механизм проведения потенциала действия.Проведение возможно только при наличии на всем протяжении или в ограниченных, но повторяющихся участках волокна потенциалзависимых ионных каналов, ответственных за формирование новых ПД. В распространении ПД можно выделить два этапа: этап распространения электрического поля, снижающего мембранный потенциал, и этап генерации новых ПД в новых участках нервного волокна. Электрическое поле - разновидность материи, посредством которой осуществляется силовое воздействие на электрические заряды, находящиеся в этом поле. Электрическое поле, которое генерируется биологическими структурами, является источником информации о состоянии клеток и органов организма. Например, состояние электрического поля сердца, записанного в виде электрокардиограммы, помогает установить возможные повреждения сердца. В зависимости от расположения и концентрации ионных каналов в мембране нервного или мышечного волокна имеются два варианта проведения ПД: непрерывный и сальтаторный (скачкообразный).
Непрерывное проведение ПД. Непрерывное проведение ПД происходит в мышечных волокнах и в безмиелиновых нервных волокнах (тип С), имеющих равномерное распределение потенциалзависимых ионных каналов по всей длине волокна, участвующих в генерации ПД. Проведение нервного импульса начинается (как и в мышечном волокне) с распространения электрического поля. Амплитуда ПД в нервном волокне (мембранный потенциал + инверсия) составляет 100- 120 мВ, постоянная длина мембраны (λm - расстояние, на котором сохраняется 37 % величины ПД в виде электрического поля) в безмиелиновых волокнах равна 0,1 - 1,0 мм. В связи с этим возникший ПД за счет своего электрического поля способен деполяризовать мембрану соседнего участка до критического уровня на расстояние от 0,1 до 1 мм.
Это означает, что на этом участке (0,1-1,0 мм) одновременно генерируются новые ПД, обусловленные движением ионов Na+ в клетку, К+ - из клетки (на распространение электрического поля время не затрачивается). Число одновременно возникающих ПД ограничивается длиной возбужденного участка - для безмиелинового волокна 0,1-1,0 мм (ПД возникают рядом друг с другом в непосредственной близости). Причем сами ПД не перемещаются (они исчезают там, где возникают). Главную роль в возникновении новых ПД играет передний ПД. Вспомогательную роль в генерации новых ПД в невозбужденных участках нервного волокна играют соседние ПД (возникшие сзади переднего ПД), так как их электрическое поле суммируется с электрическим полем переднего ПД.
Таким образом, непрерывное распространение нервного импульса, как и сальтаторное, идет через генерацию новых ПД по эстафете, когда каждый участок мембраны сначала выступает как раздражаемый электрическим полем, а затем как раздражающий (в результате формирования в нем новых ПД) (рис. 4).
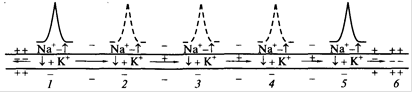
Рис. 4. Непрерывное проведение ПД по немиелинизированному нервному волокну.
Уменьшение длины горизонтальных стрелок иллюстрирует ослабление электрического поля от возбужденного участка 1. Пунктиром показан ПД, который не играет существенной роли в проведении возбуждения. 1-5 - состояние возбуждения; 6 - состояние покоя.
Сальтаторное проведение ПД. Сальтаторное проведение ПД по миелинизированным волокнам является эволюционно более поздним механизмом, возникшим впервые у позвоночных. Оно происходит в миелинизированных волокнах (типы А и В), для которых характерна концентрация потенциалзависимых ионных каналов только в небольших участках мембраны (в перехватах Ранвье), где их плотность достигает 12 000 на 1 мкм2, что примерно в 100 раз выше, чем в мембранах безмиелиновых волокон. В области миелиновых муфт, обладающих хорошими изолирующими свойствами, потенциалзависимых каналов почти нет, поэтому ПД здесь не возникают. Покрытый миелиновой муфтой участок нервного волокна между перехватами Ранвье в механизме проведения ПД выполняет роль изолятора электрического кабеля. В этих условиях ПД, возникший в одном перехвате Ранвье, за счет электрического поля деполяризует мембрану соседних перехватов до критического уровня, что приводит к возникновению в них новых ПД, т.е. возбуждение проводится скачкообразно от одних перехватов к другим (рис. 5).
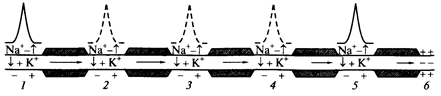

Рис. 5. Сальтаторное проведение ПД в миелинизированном нервном волокне.
Уменьшение длины горизонтальных стрелок иллюстрирует ослабление электрического поля от возбужденного участка 1. Пунктиром показаны ПД, которые не играют существенной роли в проведении возбуждения. 1-5 состояние возбуждения; 6 - состояние покоя
Na+-каналы начинают открываться при достижении деполяризации клеточной мембраны 50 % Екр. Постоянная длины мембраны миелинового волокна достигает 5 мм. Это означает, что электрическое поле ПД на данном расстоянии сохраняет 37 % своей амплитуды (около 30 мВ) и может деполяризовать мембрану до критического уровня (пороговый потенциал в перехватах Ранвье равен около 15 мВ). Благодаря этому, в случае повреждения ближайших на пути следования перехватов Ранвье ПД возбуждает 2 − 4-й и даже 5-й перехваты. Поэтому возбуждение распространяется очень быстро по всей длине волокна, а ионы движутся только перпендикулярно относительно длины волокна. Электрическое поле ПД, возникших сзади переднего ПД, суммируется с его электрическим полем, как и при непрерывном распространении возбуждения.
Возникающие ПД не могут инициировать возникновение других ПД в обратном направлении, так как нервное волокно находится еще в рефракторном состоянии. Это не противоречит тому факту, что раздражение нервного волокна в эксперименте вызывает распространение возбуждения в двух направлениях, поскольку в этом случае участки нервного волокна по обеим сторонам от места раздражения находятся в состоянии покоя. В натуральных же условиях первый ПД, инициирующий распространение возбуждения по аксону, возникает в аксонном холмике, а возбуждение проводится только в одном направлении - по аксону к другой клетке. Сравнение непрерывного и сальтаторного проведениявозбуждения показывает, что различие в механизме проведения возбуждения по миелинизированным и немиелинизированным нервным волокнам не принципиально. Оно заключается лишь в том, что очередные ПД в безмякотном волокне возникают на более близком расстоянии друг от друга, поскольку ионные каналы расположены в непосредственной близости друг от друга и непрерывно по всей длине нервного волокна. Поэтому такое проведение и назвали непрерывным. Число одновременно возникающих ПД в мякотном волокне в отличие от безмякотного строго ограничено числом возбужденных перехватов Ранвье - максимально 5.
Однако сальтаторное проведение возбуждения имеет два важных преимущества по сравнению с непрерывным проведением возбуждения.
Во-первых, сальтаторное проведение более экономично в энергетическом плане, так как возбуждаются только перехваты Ранвье, площадь которых менее 1% мембраны, и, следовательно, надо меньше энергии для восстановления трансмембранных градиентов Na+ и К+, расходующихся в процессе возникновения ПД.
Во-вторых, возбуждение в миелинизированных волокнах проводится с большей скоростью, чем в безмиелиновых волокнах, так как в них электрическое поле ПД в области миелиновых муфт распространяется значительно дальше - на соседние перехваты Ранвье, поскольку электроизоляция (миелиновые муфты) уменьшает рассеивание электрического поля. Кроме того, миелинизированные волокна в большинстве своем толще немиелинизированных, что также ускоряет проведение возбуждения, поскольку электрическое сопротивление более толстых волокон меньше.
В процессе проведения возбуждения время затрачивается только на перпендикулярное относительно мембраны волокна движение ионов в клетку и из клетки при формировании нового ПД, а влияние электрического поля возникших ПД на соседний участок распространяется вдоль длины волокна мгновенно - время распространения электрического поля практически равно нулю.
Скорость распространения возбуждения увеличивается также при большой амплитуде ПД, что является следствием формирования более сильного электрического поля, обеспечивающего критический уровень деполяризации нервного волокна на большем расстоянии.
Рассмотренные механизмы проведения ПД по нервному волокну свидетельствуют о том, что время распространения ПД в миелинизированных и немиелинизированных нервных волокнах действительно определяется только временем возникновения ПД, т. е. перпендикулярным относительно мембраны движением ионов в клетку и из клетки, поскольку электрическое поле распространяется мгновенно.
Характеристика проведения возбуждения по нервным волокнам. Нервные волокна могут проводить возбуждение в двух направлениях. Если в эксперименте нанести раздражение в любом участке нерва, то ПД будет распространяться в обе стороны от места раздражения.
Возбуждение проводится изолированно в каждом нервном волокне. Это обусловлено тем, что петли тока в межклеточной жидкости ствола, имеющей низкое сопротивление, почти не проникают в невозбужденные нервные волокна из-за большого сопротивления их оболочек. Изолированное проведение импульсов по нервным волокнам обеспечивает точное афферентное и эфферентное влияния функционально разнородных волокон нерва. Однако при одновременном раздражении большого числа нервных волокон возможно возбуждение других - прилежащих волокон и усиление нервных влияний.
Большая скорость проведения возбуждения, достигающая 120 м/с. Передача возбуждения по нервным волокнам является наиболее скоростным из известных способов передачи информации на значительные расстояния в организме.
Малая утомляемость нервного волокна.При нормальной доставке кислорода и питательных веществ проводящий возбуждение нерв практически неутомляем. Это обусловлено тем, что при проведении одного ПД по нервному волокну используется всего лишь одна миллионная часть запасов трансмембранных ионных градиентов и, следовательно, нужны небольшие количества АТФ для восстановления ионных градиентов. Расход энергии в нервном волокне на единицу массы примерно в 16 раз меньше, чем в целом организме в условиях основного обмена, и в миллион раз меньше, чем в работающей мышце.
Возможность функционального блока проведения возбуждения при морфологической целостности волокон. Нарушение физиологической непрерывности нервных волокон возникает при действии электрического тока, анестетиков, при воспалении, гипоксии, охлаждении. После прекращения действия этих факторов проведение возбуждения по нервным волокнам восстанавливается. Причиной блока проведения возбуждения является инактивация Na+-каналов.
Аксонный транспорт. Основная масса веществ, образующихся в теле нейрона (ферментов, структурных белков, полисахаридов, липидов), используется в различных его отделах. Для транспорта веществ, перечисленных медиаторов путем диффузии на расстояние, равное максимальной длине аксона (около 1 м), потребовалось бы 50 лет! Транспорт в отростках нейрона лучше изучен в аксонах и получил название аксонного транспорта. С его помощью осуществляется трофическое влияние на различные участки нейрона и на иннервируемые клетки. Транспорт веществ в дендритах осуществляется из тела клетки со скоростью около 3 мм в сутки. Различают быстрый и медленный аксонный транспорт.
Быстрый аксонный транспортидет в двух направлениях: от тела клетки до аксонных окончаний (антеградный транспорт, скорость 250-400 мм/сут) и в противоположном направлении (ретроградный транспорт, скорость 200-300 мм/сут). Посредством антеградного транспорта в окончания аксона доставляются ферменты, медиаторы, липиды, везикулы, содержащие гликопротеины мембран. При помощи ретроградного транспорта в тело нейрона переносятся везикулы, содержащие остатки разрушенных структур, фрагменты мембран, ацетилхолинэстераза, неидентифицированные «сигнальные вещества», регулирующие синтез белка в теле клетки. В патологических условиях по аксону к телу клетки могут транспортироваться вирусы бешенства, герпеса, полиомиелита, столбнячный токсин. Быстрый аксонный транспорт осуществляется с помощью специальных структурных элементов нейрона: микротрубочек и микрофиламентов. Для транспорта необходима энергия АТФ.
Медленный аксонный транспортосуществляется только в антеградном направлении и представляет собой передвижение всего столба аксоплазмы. Он выявляется в опытах со сдавливанием (перевязкой) аксона со скоростью 1 - 2 мм/сут, что соответствует скорости роста аксона в онтогенезе и при регенерации. Медленный аксонный транспорт не нарушается при разрушении микротрубочек, но прекращается при отделении аксона от тела нейрона, что свидетельствует о разных механизмах быстрого и медленного аксонного транспорта. С помощью медленного транспорта перемещаются белки микротрубочек и микрофиламентов (тубулин, актин и др.), ферменты цитозоля, РНК, белки каналов, насосов.
Аксонный транспорт играет важную роль также и при регенерации поврежденных нервных волокон.
Возбуждение от нейрона к нейрону движется по аксону и передается другому нейрону с помощью синапса.
ТЕСТ
Спинной мозг (Вариант 1)
. Нервные импульсы от рецепторов поступают:
а) в передние корешки спинного мозга
б) в задние корешки спинного мозга
в) в центральный канал спинного мозга
Ответ. [5, с.46]
Передние и задние корешки спинного мозга совершенно различны по своим функциям. Если задние корешки содержат только афферентные (чувствительные, сенсорные) нервные волокна и проводят в спинной мозг чувствительные импульсы различного характера, то передние корешки представлены только эфферентными (двигательными, или моторными, и вегетативными) волокнами, передающими нервные импульсы к эффекторам.
2. Спинномозговые нервные узлы находятся:
а) в задних корешках спинного мозга
б) в передних корешках
в) в нервных окончаниях
Ответ. [5, с.46]
В каждыйсегмент спинного мозга с обеих сторон через задние латеральные борозды входят задние (чувствительные) корешки спинномозгового нерва, которые представляют собой комплекс центральных отростков сенсорных нейронов соответствующих спинномозговых узлов. Эти узлы в количестве 31 пары обычно расположены в области межпозвоночных отверстий. Каждый их них представляет собой овальное утолщение по ходу заднего корешка и состоит из сенсорных псевдоуниполярных нейронов.
Совокупность нейронов спинномозгового узла образует ганглионарный (узловой) нервный центр, где происходит первичная обработка сенсорной (чувствительной) информации. Каждый нейрон спинномозгового узла имеет короткий отросток, сразу делящийся на два: периферический, который начинается рецепторами в коже, мышцах, суставах или внутренних органах, и центральный, направляющийся в составе заднего корешка в спинной мозг.
. Белое вещество спинного мозга проводит нервные импульсы:
а) от ЦНС к мышцам
б) от рецепторов к ЦНС
в) в обоих направлениях
Ответ. [5. с.49-51]
Снаружи от серого вещества, в котором сосредоточены тела нервных клеток, расположено белоевещество. Оно представлено длинными отростками нейронов - аксонами, покрытыми миелиновой оболочкой, придающей им белый цвет. Эти нервные волокна осуществляют связи между соседними сегментами спинного мозга, а также восходящие и нисходящие связи спинного и головного мозга.
Передние и задние борозды и щель, расположенные на поверхности спинного мозга, разделяют его белое вещество на симметрично лежащие части - канатики спинного мозга. Различают задние, боковые и передние канатики. Самую внутреннюю их часть, непосредственно прилежащие к серому веществу, составляют нервные волокна собственных пучков спинного мозга, которые осуществляют связи между соседними сегментами спинного мозга. Основная масса волокон канатиков представлена отростками тел нервных клеток, которые образуют двустороннюю связь сегментарного аппарата спинного мозга с головным мозгом. Эта связь осуществляется посредством восходящих и нисходящих проводящих путей, которые составляют белое вещество спинного мозга. По восходящим проводящим путям информация поступает из спинного мозга к головному, а по нисходящим, напротив, из головного мозга к соответствующим двигательным ядрам спинного мозга.
В формировании восходящих проводящих путей принимают участие ядра серого вещества спинного мозга, представляющие собой скопления вставочных нейронов, длинные отростки которых, направляясь к определенным отделам головного мозга, образуют соответствующие пути. Так, собственное ядро заднего рога спинного мозга связано с передачей болевой и температурной чувствительности. Через грудное и промежуточное медиальное ядра осуществляется передача в мозжечок бессознательного мышечно-суставного чувства. Отростки нервных клеток, расположенных в этих ядрах, направляются головной мозг в составе боковых канатиков.
Нисходящие проводящие пути образованы длинными отростками нейронов, лежащих в головном мозге. Эти отростки несут информацию к мотонейронам, расположенным в передних рогах спинного мозга.
В задних канатиках белого вещества спинного мозга различают два чувствительных восходящих проводящих пути: тонкий пучок, лежащий медиально, и клиновидный пучок, расположенный латерально. Оба этих пучка представляют собой скопление центральных отростков сенсорных нейронов, лежащих в спинномозговых ганглиях. Они несут непосредственно в головной мозг осознаваемую человеком сенсорную информацию от органов осязания, мышц, суставов, связок и т. д.
В боковых канатиках белого вещества спинного мозга проходит целый ряд трактов. Наиболее значимые из них.
. Двигательные нисходящие проводящие пути представляют собой совокупность отростков нейронов, расположенных в двигательных центрах головного мозга; они передают двигательные импульсы из этих центров на мотонейроны передних рогов серого вещества спинного мозга. Корково-спинномозговойлатеральный путь, или латеральный пирамидный тракт, обеспечивает проведение нервных импульсов из двигательного центра коры больших полушарий к соответствующим сегментам спинного мозга. Красноядерно-спинномозговой путь обеспечивает проведение нервных импульсов к мотонейронам соответствующих сегментов спинного мозга из подкорковых двигательных центров головного мозга. Ретикуло-спинномозговой путь проводит импульсы от ретикулярной формации головного мозга к мотонейронам в составе двигательных ядер передних рогов спинного мозга. Передача импульсов от волокон ретикуло-спинномозгового пути на мотонейроны осуществляется посредством интернейронов - клеток Реншоу.
На мотонейронах спинного мозга происходит конвергенция всех нисходящих влияний из разных отделов головного мозга, посредством которых реализуется деятельность скелетной мускулатуры.
Чувствительные (восходящие) проводящие пути бокового канатика представляют собой совокупность отростков вставочных нейронов, расположенных в ядрах серого вещества спинного мозга (в задних рогах и в промежуточном веществе). Эти пути передают сенсорные импульсы, поступающие от кожи, суставов и мышц в соответствующие центры головного мозга. Спиноталамические латеральные пути проводят импульсы болевой и температурной чувствительности в подкорковые центры общей чувствительности (таламусы), расположенные в области промежуточного мозга, и далее - к соматосенсорной коре больших полушарий. Спинно-мозжечковые пути (передний и задний) несут в мозжечок импульсы бессознательного мышечно-суставного (проприоцептивного) чувства.
В передних канатиках белого вещества спинного мозга среди нисходящих расположен двигательный проводящий путь - передний корково-спинномозговой путь (передний пирамидный тракт). Этот путь подобно латеральному корково-спинномозговому пути состоит из нервных волокон, являющихся отростками пирамидных нейронов двигательного центра коры и проводит двигательные импульсы к мотонейронам соответствующих сегментов серого вещества спинного мозга. Среди восходящих проводящих путей следует отметить передний спинно-таламический путь, связанный с передачей в подкорковые чувствительные центры (таламусы) чувства осязания и давления.
4. К спинномозговым рефлексам млекопитающих животных относится:
а) мочеиспускание
б) охрана потомства
в) строительство нор
Ответ. [1, с.61-63]
Спинной мозг является центром ряда рефлексов. Основные - это проприоцептивные рефлексы: моно-, би- и полисинаптические сухожильные и миостатические, хотя немалую роль играют и кожные, и висцеральные. Некоторые спинномозговые рефлексы имеют характер ритмических (чесательный, шагательный и др.), в основе которых лежат реципрокные отношения в спинальных мотонейронных пулах и феномен отдачи. Мышечные рефлексы обеспечивают быстрые движения (фазные рефлексы) и изменение и поддержание позы (тонические); эти рефлексы могут возникать от проприо- и экстероцепторов кожи, поддерживаются же они благодаря связи от проприоцепторов.
В ряде случаев спинальные рефлексы протекают настолько константно, что изменение и выпадение их может являться диагностическим тестом состояния спинного мозга. В спинном мозгу локализован также ряд эффекторных центров, осуществляющих вегетативные функции - сосудодвигательные, потоотделительные, мочеполовые, дефекационные и др.
Таким образом, спинной мозг, являясь главным исполнительным отделом центральной нервной системы, в то же время участвует в первичной обработке проприо- висцеро- и экстероцептивных кожных сигналов, осуществляет координационно-интегративную рефлекторную функцию на сегментарном уровне и обеспечивает обратную афферентацию от проприоцептивного аппарата к управляющим структурам головного мозга.
Охрана потомства, строительство нор к спинномозговым рефлексам млекопитающих животных не относится.
5. Какова функция вставочных нейронов спинного мозга?
а) восприятие раздражения
б) осуществление связи с отделами ЦНС
в) проведение возбуждения к мышцам
Ответ. [5, с.46-49]
На поперечном срезе спинного мозга видно, что его вещество неоднородно. Внутри расположено серое вещество, а снаружи - белое вещество. Серое вещество спинного мозга представляет собой скопление тел нейронов и их коротких отростков, белое вещество - скопление их длинных отростков, соединяющих нервные клетки различных сегментов спинного мозга между собой и с клетками головного мозга. В центре серого вещества имеется центральный канал, по которому циркулирует спинномозговая жидкость.
В сером веществе спинного мозга различают задние, промежуточные и передние столбы; на поперечном срезе они выглядят как передние и задние рога. В грудном отделе спинного мозга в сером веществе выделяют еще боковые рога. В задних рогах и в промежуточном веществе группируются вставочные нейроны (интернейроны), а в передних рогах - мотонейроны.
Локальные скопления нервных клеток в сером веществе называют ядрами. В ядрах осуществляется обработка поступающей в спинной мозг информации и передача ее на другие нервные центры.
Поступающие в составе задних корешков нервные волокна, являющиеся отростками сенсорных нейронов, лежащих в спинномозговых узлах, частично заканчиваются в области верхушки заднего рога, которая носит название студенистого вещества. Студенистое вещество, или желатинозная субстанция, представляет собой скопление интернейронов (пучковых клеток), выполняющих в рефлекторной дуге роль промежуточного, вставочного звена между сенсорными нейронами и мотонейронами. Пучковые клетки в большом количестве встречаются не только в задних рогах, но и в промежуточном веществе. Благодаря своим отросткам они осуществляют связь выше- и нижележащих сегментов спинного мозга между собой. Аксоны этих вставочных нейронов достигают передних рогов спинного мозга, где заканчиваются на мотонейронах, иннервирующих соответствующие скелетные мышцы.
Таким образом, основная функция сегментарного аппарата спинного мозга, в состав которого входит участок серого вещества вместе с соответствующей парой спинномозговых нервов и относящихся к ним передних и задних корешков, сводится к осуществлению врожденных сегментарных рефлексов.
В задних рогах (столбах) нейроны располагаются послойно. Здесь сосредоточено множество ядер, на которых оканчиваются афферентные нервные волокна общей соматической и висцеральной чувствительности. Среди ядер заднего рога следует отметить собственное ядро, которое располагается в центре заднего рога. Оно служит местом переключения тех чувствительных нервных волокон, которые несут информацию о болевых и температурных раздражениях с поверхности кожи, и передачи этой информации по восходящим трактам в головной мозг.
В промежуточных столбах лежат: грудное ядро (ядро Кларка), промежуточное медиальное ядро и промежуточное латеральное ядро. Последнее расположено в боковом роге и прослеживается только в грудных сегментах спинного мозга. Клетки промежуточного латерального ядра связаны с рефлекторной деятельностью внутренних органов и составляют центры вегетативной нервной системы. Грудное ядро и промежуточное медиальное ядро имеют отношение к переключению афферентной соматической информации, поступающей в спинной мозг по чувствительным нервным волокнам в составе задних корешков, на проводящие пути, ведущие к головному мозгу, в частности к мозжечку. В передних рогах (столбах) серого вещества лежат моторные (двигательные) ядра. В местах шейного и поясничного утолщений спинного мозга, от сегментов которых осуществляется иннервация верхней и нижней конечностей, выделяют до пяти групп двигательных ядер. В двигательных ядрах лежат мотонейроны, аксоны которых в составе соответствующих нервов достигают мышечных волокон и передают им необходимые команды из центральной системы.
ОТВЕТИТЬ НА ВОПРОСЫ
. Утолщения спинного мозга появляются в шейном и поясничном отделах. Почему? С чем это связано?
Ответ. [8, с. 146]
Спинной мозг представляет собой тяж длиной около 45 см у мужчин и около 42 см - у женщин, имеет сегментарное строение (31-33 сегмента); каждый его сегмент связан с определенной частью тела. Спинной мозг включает пять отделов: шейный, грудной, поясничный, крестцовый и копчиковый.
В процессе эволюции сформировалось два утолщения: шейное (сегменты, иннервирующие верхние конечности) и пояснично-крестцовое (сегменты, иннервирующие нижние конечности) как результат повышенной нагрузки на эти отделы спинного мозга. Тренировка любого органа обеспечивает прогрессивное его развитие, естественно, при этом совершенствуется и функция. Орган, который не получает достаточной нагрузки, постепенно атрофируется. В указанных утолщениях спинного мозга соматические нейроны наиболее крупные, их больше, в каждом корешке этих сегментов больше нервных волокон, они имеют наибольшую толщину.
. Какое значение имеют восходящие и нисходящие пути спинного мозга?
Ответ. [8, стр.148-150]
Проводниковая функция осуществляется с помощью нисходящих и восходящих путей. Афферентная информация поступает в спинной мозг через задние корешки, эфферентная импульсация и регуляция функций различных органов и тканей организма осуществляется через передние корешки. Каждый корешок представляет собой множество нервных волокон.
Все афферентные входы в спинной мозг несут информацию от трех групп рецепторов: 1) от кожных рецепторов (болевых, температурных, прикосновения, давления, вибрации); 2) от проприорецепторов (мышечных ― мышечных веретен, сухожильных - рецепторов Гольджи, надкостницы и оболочек суставов); 3) от рецепторов внутренних органов - висцерорецепторов (механо- и хеморецепторов).
Значение афферентной импульсации, поступающей в спинной мозг, заключается в следующем: 1) участие в координационной деятельности ЦНС по управлению скелетной мускулатурой; при выключении афферентной импульсации от рабочего органа управление им становится несовершенным; 2) участие в процессах регуляции функций внутренних органов; 3) поддерживает тонус ЦНС; при выключении афферентной импульсации наступает уменьшение суммарной тонической активности ЦНС; 4) несет информацию об изменениях окружающей среды. Основные проводящие пути спинного мозга приведены в Табл. 1.
Таблица 1
| Восходящие (чувствительные) пути | Физиологическое значение |
| Клиновидный пучок (пучок Бурдаха) проходит в задних столбах, импульсация поступает в кору | Осознаваемая проприорецептивная импульсация от нижней части туловища и ног |
| Тонкий пучок (пучок Голля), проходит в задних столбах, импульсация поступает в кору | Осознаваемая проприорецептивная импульсация от верхней части туловища и рук |
| Задний спинно-мозжечковый (Флексига) | Не осознаваемая проприорецептивная импульсация |
| Передний спинно-мозжечковый (Говерса) | То же |
| Латеральный спиноталамический | Болевая и температурная чувствительность |
| Передний спиноталамический | Тактильная чувствительность, прикосновение, давление |
| Нисходящие (двигательные) пути | Физиологическое значение |
| Латеральный кортикоспинальный (пирамидный) | Импульсы к скелетным мышцам, произвольные движения |
| Передний кортикоспинальный (пирамидный) | То же |
| Руброспинальный (Монакова), проходит в боковых столбах | Импульсы, поддерживающие тонус скелетных мышц |
| Ретикулоспинальный, проходит в передних столбах | Импульсы, поддерживающие тонус скелетных мышц с помощью возбуждающих и тормозящих влияний на a- и y-мотонейроны, а также регулирующие состояние спинальных вегетативных центров |
| Вестибулоспинальный, проходит в передних столбах | Импульсы, обеспечивающие поддержание позы и равновесия тела |
| Тектоспинальный, проходит в передних столбах | Импульсы, обеспечивающие осуществление зрительных и слуховых двигательных рефлексов (рефлексов четверохолмия) |
РЕШИТЬ ЗАДАЧИ
У некоторых пациентов коленный рефлекс бывает слабо выражен. Чтобы усилить его, предлагают сцепить руки и тянуть их в разные стороны. Почему это приводит к усилению рефлекса?
Ответ. [8, с. 273-276]
Это связано, по-видимому, с тем, что сцепить руки и растягивая их в разные стороны, пациент возбуждает нейроны, длинные аксоны которых входят в состав спинноталамических путей. При этом, от болевых рецепторов туловища, шеи и конечностей Аδ- и С-волокна первых чувствительных нейронов (их тела находятся в спинальных ганглиях) идут в составе спинномозговых нервов и входят через задние корешки в спинной мозг, где разветвляются в задних столбах и образуют синаптические связи прямо или через интернейроны со вторыми чувствительными нейронами, длинные аксоны которых входят в состав спиноталамических путей. При этом они возбуждают два вида нейронов: одни нейроны активируются только болевыми стимулами, другие - конвергентные нейроны - возбуждаются также и неболевыми стимулами. Вторые нейроны болевой чувствительности преимущественно входят в состав боковых спиноталамических путей, которые и проводят большую часть болевых импульсов. На уровне спинного мозга аксоны этих нейронов переходят на сторону, противоположную раздражению, в стволе головного мозга они доходят до таламуса и образуют синапсы на нейронах его ядер. Часть болевой импульсации первых афферентных нейронов переключается через интернейроны на мотонейроны мышц-сгибателей и участвуют в формировании защитных болевых рефлексов.
В качестве дополнения можно добавить, что болевые рецепторы (ноцицепторы) являются свободными окончаниями чувствительных миелиновых нервных волокон Аδ и немиелиновых волокон С. Они найдены в коже, слизистых оболочках, надкостнице, зубах, мышцах, суставах, внутренних органах и их оболочках, сосудах и других участках тела; они не обнаружены в нервной ткани головного мозга и костном мозге.
Выделяют следующие основные типы болевых рецепторов. 1. Механоноцицепторы и механотермические ноцицепторы Аδ -волокон реагируют на сильные механические и термические раздражители, проводят быструю механическую и термическую боль, быстро адаптируются; они расположены преимущественно в коже, мышцах, суставах, надкостнице; их афферентные нейроны имеют малые рецептивньие поля. 2. Полисенсорные ноцицепторы С-волокон реагируют на механические, термические (тепловой и холодовой) и химические раздражители, проводят позднюю плохо локализованную боль, медленно адаптируются.
Болевые рецепторы возбуждаются тремя видами раздражителей: 1) механические раздражители, создающие давление при сдавливании, растяжении, сгибании, скручивании; 2) термические раздражители (тепловыми - свыше 45°С и холодовыми - ниже 15°С); 3) химические раздражители. Болевые рецепторы имеют высокий порог возбуждения (низкую возбудимость), что обеспечивает их ответ только на чрезвычайные раздражители. Ноцицепторы С-афферентов плохо адаптируются к длительно действующим раздражителям. Возможно повышение чувствительности болевых рецепторов - снижение порога их раздражения при многократной или длительной стимуляции, что называется гипералгезией. При этом ноцицепторы способны отвечать на стимулы субпороговой величины, а также возбуждаться раздражителями других модальностей.
У больного обнаружены нарушения деятельности сердечно-сосудистой системы, у другого больного - нарушения деятельности желудочно-кишечного тракта. Консилиум врачей направил их на лечение не в терапевтическую клинику, а в неврологическую. Чем продиктовано такое решение?
Ответ. Сложно оценить действия врачей, не имея на руках ни результатов клинического обследования больных, ни результатов их анализов. Вероятнее всего, решение врачей направить этих больных в неврологическую клинику объясняется тем, что в результате обследова