Групп 127
Фадеев С.В.
Текст для презентации
«ДНК»
Цель модуля – ознакомление с историей открытия нуклеиновых кислот, уровнями структурной организации и функциями ДНК; молекулярным строением и функциями различных видов РНК
СТРАНИЦА
Нуклеиновые кислоты были открыты в 1868 г. швейцарским биологом Фридрихом Мишером. Он выделил из ядер клеток гноя вещество, которое назвал нуклеином (от лат. «nucleus» - ядро). Клетки гноя – это лейкоциты. Мишер обнаружил, что нуклеин содержит кислый компонент, который известен теперь как ДНК. Мишер выделил фракцию нуклеиновой кислоты и изучал ее свойства. Однако структура ДНК, оставалась невыясненной до конца 40-х годов.
СТРАНИЦА
В настоящее время установлено, что ДНК – это чрезвычайно длинные полимерные цепи. Их молекулярный вес от нескольких до сотен тысяч миллионов. ДНК обычно состоит из 2-х цепей. Цепи ДНК состоят из соединенных друг с другом мономерных единиц – дезоксирибонуклеотидов четырех разных типов.
Нуклеотид состоит из 3 компонентов:
1. пуриновое или пиримидиновое основание;
2. моносахарид: рибоза (в РНК) или дезоксирибоза (в ДНК);
3. остаток фосфорной кислоты.
Нуклеиновые кислоты имеют различающийся состав. В частности, дезоксирибонуклеиновые кислоты (ДНК) содержат дезоксирибозу, а рибонуклеиновые кислоты (РНК) - рибозу. Эти и другие отличия в составе нуклеиновых кислот приведены ниже:
Уровни структурной организации ДНК
Нуклеозиды - соединения азотистого основания и углеводов (рибозы и дезоксирибозы). Нуклеозиды образуются за счет N-гликозидной связи между ДЕВЯТЫМ атомом азота у ПУРИНОВЫХ (первым атомом азота- у ПИРИМИДИНОВЫХ) оснований и гидроксилом ПЕРВОГО атома углерода рибозы или дезоксирибозы (таблица 1). Во избежание путаницы, нумерация атомов азотистых оснований осуществляется арабскими цифрами, а у атомов углерода рибоз - арабскими цифрами со “штрихом”.
Таблица 1 Нуклеозиды
| Пурино-вые | 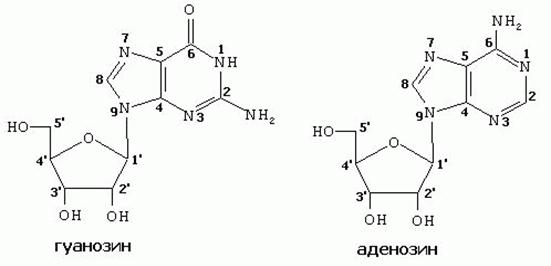
|
| Пирими-ди-новые | 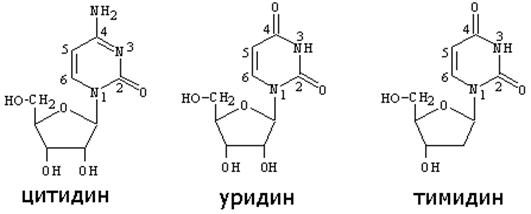
|
Нуклеотиды отличаются от нуклеозидов наличием остатков фосфорной кислоты (от одного до трех), связанных простой эфирной связью с гидроксилом 5’ атома углерода рибоз. Остатки фосфорных кислот между собой также связаны простой эфирной связью. В зависимости от числа остатков фосфорной кислоты в нуклеотидах различают МОНО-, ДИ- и ТРИФОСФОНУКЛЕОТИДЫ. Их номенклатура приведена ниже:
| азотистые основания | нуклеозиды | Нуклеотиды | |
| полное название | Сокращенное | ||
| аденин | аденозин | Аденозинмонофосфат | АМФ |
| гуанин | гуанозин | Гуанозинмонофосфат | ГМФ |
| цитозин | цитидин | Цитидинмонофосфат | ЦМФ |
| урацил | уридин | Уридинмонофосфат | УМФ |
| тимин | тимидин | тимидинмонофосфат | ТМФ |
ДНК и РНК обладают первичной, вторичной, третичной структурами.
Первичная структура ДНК – последовательность нуклеотидов в молекуле ДНК.
Нуклеотиды связываются между собой за счет остатков фосфорной кислоты.
Нуклеотидные единицы соединяются между собой фосфодиэфирными мостиками. Фосфодиэфирные мостики соединяют 3¢ - гидроксильную группу одного нуклеотида с 5¢ - гидроксильной группой другого.
Поскольку основой нуклеиновых кислот является САХАРОФОСФАТНЫЙ ОСТОВ, в сокращенных написаниях участков цепи используют лишь ОДНОБУКВЕННЫЕ СИМВОЛЫсоответствующего азотистого основания. Полное и схематичное обозначение участка полинуклеотидной цепи приведены ниже:
5’-НО-Г-A-A- T-Ц-T-A-Ц-A-…3'
Вследствие наличия сильно-диссоциирующих фосфатных групп, нуклеиновые кислоты (НК) легко образуют связи с основными белками с образованием НУКЛЕОПРОТЕИНОВ. Протеины отделяются от НК детергентами. После расщепления белков протеиназами НК осаждаются спиртом.
В ДНК содержатся минорные основания обычно – это метилированные формы главных оснований, например, 5-метилцитозин и 6-метиладенин. Метилирование оснований происходит уже после репликации ДНК. Эти метилированные основания защищают «свои» ДНК от расщепления ферментами – ДНКазами. В ряде вирусных ДНК некоторые основания могут быть гидроксиметилированы или глюкозилированы. Такие минорные основания в ДНК служат специфическими сигналами, которые играют важную роль в реализации генетической информации или в обеспечении ее сохранности. Минорные основания были обнаружены также в РНК, в основном в т-РНК.
Итак, ДНК была открыта в клеточном ядре в 1868 году, но ее генетическая функция оставалась невыясненной до 1943.
В 1943 г. Эвери и его сотрудники из Рокфеллеровского института обнаружили, что непатогенный (неболезнетворный) штамм бактерии пневмококка может быть трансформирован в патогенной простым добавлением ДНК, выделенной из патогенных пневмококков (свойство патогенности наследуется).
Вывод: ДНК может содержать генетическую информацию.
Было установлено, что:
1. Содержание ДНК в любой клетке или организме строго постоянно;
2. Содержание ДНК в клетке увеличивается с возрастанием сложности клетки, следовательно, с возрастанием генетической информации в клетке. Чем сложнее организм, тем больше ДНК в его клетках;
3. Гаплоидные половые клетки высших организмов (содержащие одинарный набор хромосом) содержат точно половину количества ДНК, которое обнаружено в диплоидных клетках того же организма;
4. ДНК-содержащие вирусы бактерий (бактериофаги) и вирусы животных, имеющие лишь несколько генов, содержат очень мало ДНК.
Это указывает на генетическую функцию ДНК: ДНК хранит генетическую информацию.
В настоящее время установлено, что функция ДНК состоит в том, что она хранит генетическую информацию, необходимую для кодирования структуры всех белков и всех РНК каждого вида организма, регулирует во времени и пространстве биосинтез компонентов клеток и тканей, определяет деятельность организма в течение его жизненного цикла и обеспечивает индивидуальность данного организма.
Самым важным ключом к разгадке структуры ДНК стало открытие, сделанное в конце 40-х годов Чаргаффом и его коллегами из Колумбийского университета. Они обнаружили, что 4 основания встречаются в ДНК разных организмов в различных соотношениях и что между основаниями существует определенная количественная связь. Такого рода результаты, полученные для препаратов ДНК, выделенных из огромного множества различных видов, привели Чаргаффа и более поздних исследователей к следующим выводам.
Вторичная структура ДНК
Правила Чаргаффа
1. Препараты ДНК, выделенные из разных тканей одного и того же организма, имеют одинаковый нуклеотидный состав.
2. Нуклеотидный состав ДНК у разных видов различен.
3. Число адениновых остатков в любой ДНК независимо от вида организма равно числу тиминовых остатков (А=Т), а число гуаниновых остатков всегда равно числу цитозиновых остатков (Г=Ц).
Следовательно, сумма пуриновых остатков равно сумме пиримидиновых, т.е. А + Г = Т + Ц
Было установлено, что ДНК близких видов имеют сходный нуклеотидный состав, а эволюционно отдаленные организмы заметно отличаются по нуклеотидному составу.
Молекула ДНК компактна. Компактность создает вторичная и третичная структура ДНК.
В 1953 году американский генетик Джеймс Уотсон и английский физик Френсис Крик как результат своих работ в Кембриджском университете (Англия) предложили трехмерную модель двойной спирали ДНК. Их модель была основана на данных рентгеноструктурного анализа ДНК и объясняла характерную для ДНК парность оснований. Модель состоит из двух цепей ДНК, закрученных в спираль вправо вокруг одной и той же оси с образованием двойной спирали. Две цепи в этой спирали антипараллельны, т.е. их 5¢, 3¢ - межнуклеотидные фосфороэфирные мостики направлены в противоположные стороны
5¢ 3’

3¢ 5’
Гидрофильные остовы цепей, состоящие из чередующихся остатков дезоксирибозы и отрицательно заряженных фосфатных групп, расположены на внешней стороне двойной спирали и обращены в сторону окружающей ее воды. Гидрофобные пуриновые и пиримидиновые основания обеих цепей уложены стопкой внутри двойной спирали, так что практически плоские молекулы оснований сближены между собой и расположены перпендикулярно длинной оси двойной спирали. Пространственное взаиморасположение цепей приводят к возникновению большой и малой бороздок. Основания одной цепи спарены с находящимися в той же плоскости основаниями другой цепи, т.е. две цепи в молекуле ДНК стабилизированы водородными связями: между А и Т возникают две водородные связи (А = Т), а между Г и Ц три водородные связи (Г = Ц).
С помощью данных рентгеноструктурного анализа установлено, что:
1. Молекула ДНК имеет постоянный диаметр 2 нм (1 нм =10-9м)
молекула пурина 1,2 нм
молекула пиримидина 0,8 нм
1,2 + 0,8 = 2 нм
Количество пуриновых оснований равно количеству пиримидиновых оснований.
2. Основания уложенных стопкой внутри двойной спирали ДНК и располагаются на расстояниях 0,34 нм друг от друга.
3. На каждый полный оборот двойной спирали приходится 10 пар оснований. Следовательно, молекула ДНК имеет периодичность в структуре равную 3,4 нм.
Стабилизация двойной спирали происходит:
1. за счет водородных связей между комплементарными основаниями
А = Т, Г = Ц.
2. за счет гидрофобных взаимодействий между основаниями, благодаря чему основания оказываются спрятанными внутрь двойной спирали и защищены от соприкосновения с водой.
Таким образом, вторичная структура ДНК представляет собой свернутые в спираль две комплементарно взаимодействующие и антипараллельные полинуклеотидные цепи. Образование вторичной структуры нуклеиновых кислот возможно вследствие проявления эффектов комплементарности и стэкинг-взаимодействий.
Комплементарность - последовательность нуклеотидов в одной цепи автоматически определяет строго соответствующую ей последовательность нуклеотидов в КОМПЛЕМЕНТАРНОЙ ей цепи. Так, азотистое основание Аденин (А) всегда взаимодействует только с комплементарным ему азотистым основанием Тимин (Т) в молекулах ДНК. Одновременно азотистое основание Гуанин (Г) одной цепи взаимодействует только с комплементарным азотистым основанием Цитозин (Ц) другой цепи (как в ДНК, так и в РНК). Комплементарность оснований обеспечивается системой водородных связей. В молекулах РНК, имеющих, в основном, однонитевую структуру, на отдельных участках, азотистые основания А взаимодействуют с комплементарными им азотистыми основаниями Урацил (У).
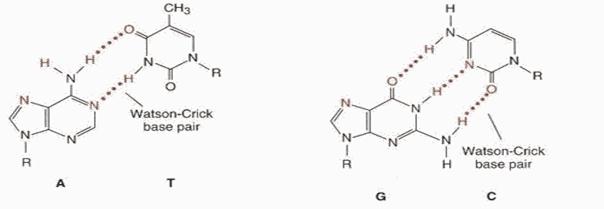
Аналогично происходит взаимодействие в процессах транскрипции, когда на матрице ДНК синтезируется молекула РНК (матричная, транспортная и т.д.), и наоборот, когда при участии РЕВЕРС-ТРАНСКРИПТАЗЫпроисходит синтез кДНК на матрице РНК.
Стэкинг-взаимодействия - особого рода (Ван-дер-Ваальсовы) взаимодействия между выложенными в стопку (как монеты) друг над другом азотистыми основаниями.
Имеются А, В, С и Z-формы ДНК, отличающиеся диаметром и шагом спирали, числом пар оснований в витке, углом наклона плоскости азотистых оснований по отношению к оси молекулы (рис. 1).
В форме В, описанной моделью Дж.Уотсона и Ф.Крика, на 1 виток спирали приходится 10 пар оснований, шаг спирали 3,4 нм, диаметр 1,8 нм, угол наклона к оси 0º. Форма В, по-видимому, благоприятна для процесса репликации. В форме А на 1 виток приходится 11 пар оснований, шаг спирали 2,8 нм, угол наклона плоскости оснований к оси составляет 20º. Форма А является предпочтительной для процессов транскрипции. Форма С, выявленная у ряда вирусов и в составе надмолекулярных структур хроматина, имеет 9,3 пары оснований в витке с углом наклона 5º. Z-форма ДНК – наименее скрученная (12 пар оснований на виток). Она представляет собой левозакрученную двойную спираль, в которой фосфоэфирный остов расположен зигзагообразно вдоль оси. Z-форма обладает только одной бороздкой. Z-форма возникает, как правило, при высоких концентрациях солей, спермина, спермидина, при метилировании остатков дезоксицитидина, при высоком содержании отрицательных супервитков в молекуле ДНК, может участвовать в регуляции экспрессии генов. Описанные формы ДНК способны к взаимно обратимым переходам в зависимости от условий среды.
В желобах вдоль всей молекулы ДНК располагаются белки: гистоны, негистоновые. Гистоны – это небольшие щелочные белки. Преобладают в хроматине соматических клеток эукариот. В них преобладают аминокислоты аргинин, лизин (до
25 %), имеющие положительный заряд на радикале. Существует 5 основных фракций гистонов Н1, Н2А, Н2В, Н3, Н4. Они отличаются содержанием аргинина и лизина.
Радикалы аминокислот, входящих в состав гистонов, могут быть метилированы, фосфорилированы или ацетилированы.
Так как гистоновые белки положительно заряжены, а ДНК имеет отрицательный заряд (из-за остатков фосфорной кислоты), то образуется ДНК-гистоновый комплекс. Гистоны образуют с молекулами ДНК прочные электростатические связи.
Негистоновые белки – это кислые белки, в них много глутаминовой и аспарагиновой аминокислот. Белки находятся в большой и малой бороздке молекулы ДНК
ДНК прокариот
Третичная структура ДНК
Различают следующие формы ДНК: ЛИНЕЙНАЯ, КОЛЬЦЕВАЯ, одно- и двух-ЦЕПОЧЕЧНАЯ (рис. 2). Двуцепочечные ДНК с "липкими" концами могут образовывать кольцо, которое далее ковалентно сшивается по сахарофосфатной цепи при помощи ДНК-лигазы.
В прокариотических клетках ДНК представляет собой замкнутое двухцепочечное кольцо. ДНК является сверхскрученной – сверхспирализованной. Суперспирализация создает третичную структуру ДНК. Сверхспиральная ДНК более компактна, чем раскрученное кольцо. Анализ ДНК, выделенной из Е.coli, показал, что она образует множество петель, поддерживаемых вместе с помощью белков. Каждая из этих петель в свою очередь сверхспирализована. Образование петель и сверхспиральность помогает обеспечить упаковку очень больших кольцевых молекул ДНК в малых объемах. Например, ДНК Е.coli имеет длину более 1 мм, в то время как длина клетки не превышает 5 мкм.
Кроме очень большой кольцевой хромосомы, расположенной в ядерной зоне, большинство бактерий содержит одну или несколько небольших кольцевых молекул ДНК, которые находятся в свободном состоянии в цитоплазме клетки. Это плазмиды. Они малы и содержат лишь несколько генов (в хромосоме тысячи генов). Некоторые плазмиды несут гены, которые обеспечивают устойчивость бактерии к антибиотикам (например, к тетрациклину, стрептомицину). Плазмиды могут переноситься из устойчивых к антибиотику клеток в чувствительные клетки того же или другого вида, делая эти клетки устойчивыми.
Плазмиды можно легко выделить из бактериальных клеток. В выделенную плазмиду встраивают новые гены из других видов и такую модифицированную плазмиду вводят обратно в обычную для нее клетку-хозяина. Плазмида, несущая чужеродный ген будет реплицироваться (удваиваться с образованием дочерних плазмид) и транскрибироваться и может заставить клетку-хозяина синтезировать белки, кодируемые искусственно встроенным геном. Молекула ДНК с искусственно встроенным геном называется рекомбинантной. С помощью рекомбинантных ДНК сейчас получают белковый гормон – инсулин, защитный белок интерферон и др.
Эукариоты имеют оформленное ядро, содержащее ДНК. Размер типичной эукариотической клетки, например, клетки печени человека, составляет в поперечнике ~ 25 мкм. Ее ядро, размером ~ 5 мкм в диаметре, содержит 46 хромосом, суммарная длина ДНК которых равна 2 м. Эукариоты содержат значительно больше ДНК, чем прокариоты. Так, клетки человека и других млекопитающих содержат в 600 раз больше ДНК, чем Е.coli. Общая длина всей ДНК, выделенной из клеток организма взрослого человека, составляет ~ 2 х 1013 м или 2 х 1010 км, что превышает окружность земного шара (4 х 104 км) и расстояние от Земли до Солнца (1,44 х 108 км).
У эукариот ДНК находится в хромосомах. В клетках человека 46 хромосом (хроматид), которые организованы в 23 пары. Каждая хромосома эукариотической клетки содержит одну очень большую молекулу двухцепочечной ДНК, несущую набор генов. Совокупность генов клетки составляет ее геном. Гены – это участки ДНК, которые кодируют полипептидные цепи и РНК.
Молекулы ДНК в 46 хромосомах человека не одинаковы по размеру. Средняя длина хромосомы составляет 130 млн. пар оснований и имеет длину 5 см. Понятно, что уместить такую ДНК в ядре возможно только путем ее определенной упаковки. При образовании третичной структуры ДНК человека происходит в среднем уменьшение ее размеров в 100 тысяч раз.
Упаковка ДНК в эукариотических хромосомах отличается от ее упаковки в прокариотических хромосомах. Эукариотические ДНК имеют не кольцевую, а линейную двухцепочечную структуру. Кроме того, третичная структура ДНК у эукариотических клеток отличается тем, что многократная спирализация ДНК сопровождается образованием комплексов с белками. ДНК эукариот содержит экзоны - участки, кодирующие полипептидные цепи, и интроны – некодирующие участки (выполняют регуляторную функцию).
Эукариотические хромосомы состоят из хроматиновых волокон. Эукариотические хромосомы выглядят как резко очерченные структуры только непосредственно до и во время митоза- процесса деления ядра в соматических клетках. В покоящихся, неделящихся эукариотических клетках хромосомный материал, называемый хроматином, выглядит нечетко и как бы беспорядочно распределен по всему ядру. Однако, когда клетка готовится к делению, хроматин уплотняется и собирается в хромосомы.
Хроматин состоит из очень тонких волокон, которые содержат ~ 60 % белка, ~ 35 % ДНК, и, вероятно, ~ 5 % РНК. Хроматиновые волокна в хромосоме свернуты и образуют множество узелков и петель. ДНК в хроматине прочно связана с белками-гистонами, функция которых состоит в упаковке и упорядочении ДНК в структурные единицы – нуклеосомы. В хроматине содержится также ряд негистоновых белков. Хроматиновые волокна напоминают по внешнему виду нитки бус. Бусинки – это нуклеосомы.
Нуклеосома состоит из белков-гистонов. Каждая нуклеосома содержит 8 молекул гистонов – по 2 молекулы Н2А. Н2В, Н3, Н4. Двухцепочечная ДНК обвивает нуклеосому дважды (рис. 3).
Нить ДНК намотана на гистоновое ядро нуклеосомы снаружи. В промежутках между нуклеосомами расположена соединительная нить ДНК, с которой связывается гистон Н1. Таким образом, нуклеосомы – это структурные единицы хроматина, выполняют функцию плотной упаковки ДНК. ДНК укорачивается за счет того, что она обвивает гистоны. Хроматин связан также с негистоновыми белками ядра, которые образуют ядерный матрикс (рис. 4).
Эукариотические клетки содержат также цитоплазматическую ДНК.
Кроме ДНК в ядре у эукариот есть ДНК в митохондриях. Хлоропласты фотосинтезирующих клеток также содержат ДНК. Обычно ДНК в цитоплазме составляет » 0, 1 % всей клеточной ДНК.
Митохондриальные ДНК – это двухцепочечные кольцевые молекулы малого размера. Молекулы ДНК в хлоропластах значительно больше, чем в митохондриях. ДНК митохондрий и хлоропластов не связана с гистонами.