Теория парения.
Первым, кто пытался объяснить сезонные и локальные изменения формы у планктических организмов, был Везенберг-Лунд (Wesenberg-Lund, 1900). В результате обстоятельных исследований Везенберг-Лунд пришел к убеждению, что решающее значение имеют здесь внешние факторы. Уже то обстоятельство, что сезонные изменения возникают скачкообразно, в течение каких-то трех недель в первой половине лета, давало автору основание искать причину их именно во внешних условиях. Еще более в пользу такого убеждения говорило то, что сезонные вариации не ограничены одними кладоцерами, но встречаются и у представителей других групп и притом у таких, которые живут пелагически, причем у всех сочленов пелагического сообщества эти изменения наступают одновременно и в одно и то же время образуют самые крайние формы.
Далее, так как изменения формы всегда начинаются при температурах 12—16° C и исчезают при почти сходных температурах и так как на Крайнем Севере и на высоких горах, где в озерах такие температуры никогда не достигаются, изменения формы отсутствуют, то Везенберг-Лунд считал вполне естественным искать решающий фактор в происходящих одновременно с изменением формы изменениях в условиях парения.
Поскольку наблюдения на датских озерах показали, что формы с увеличенной поверхностью и с выростами на теле появляются только тогда, когда вода нагревается и, следовательно, имеет меньший удельный вес, и достигают наивысшего своего выражения при максимальной температуре воды, то Везенберг-Лунд такое увеличение поверхности рассматривал как приспособление к ухудшенным в нагретой воде условиям парения.
Когда же Оствальд (Ostwald, 1902) в результате своих экспериментальных исследований показал, что гораздо большее значение, чем изменение удельного веса воды, должно иметь изменение в связи с изменением температуры и вязкости, или внутреннего трения, воды, то Везенберг-Лунд позже (Wesenberg- Lund, 1939) по существу присоединился к этому заключению Оствальда.
В самом деле, если в воде при 25°С вязкость воды уменьшается вдвое по сравнению с тем, что имеет место в воде при 0° С, то это означает, что организм, для того чтобы замедлить свое погружение и держаться во взвешенном состоянии в том же слое воды в летней воде, должен затратить двойную энергию по сравнению с затратой энергии зимой.
Так как все изменения формы направлены к тому, чтобы увеличить летним животным их относительную поверхность путем уменьшения их размеров, путем удлинения их продольной оси или образования разнообразных выростов, понижающих скорость их погружения, то естественно, сезонные вариации должны рассматриваться как приспособления к меняющейся вязкости воды.
Такое толкование сезонных вариаций казалось весьма простым и убедительным. В пользу него, кроме факта отсутствия сезонных вариаций на Крайнем Севере и в холодных альпийских озерах, говорила также слабая выраженность сезонных вариаций у всех донных и береговых форм.
Кроме того, теория парения получала себе поддержку в наличии сезонных вариаций у круглогодичных форм и в отсутствии вариаций у таких форм, которые живут в пелагиали исключительно в летнюю половину года при температуре, изменяющейся в интервале 15—30° C, тогда как в остальное время года они выпадают из планктона, образуя покоящиеся яйца, перезимовывающие на дне озер или у берега.
Эти исключительно летние животные отличаются друг от друга формой тела: значительным удлинением продольной оси тела (Leptodora), очень сильным развитием второй пары антенн (Diaphanosoma), образованием типов (Bythotrephes), мощным развитием студенистой оболочки (Holopediam), т. е. такими особенностями строения, которые находятся в хорошем соответствии с особенностями организации летних рас видов круглогодичных, встречающихся в водоемах как летом, когда скорость опускания велика, так и зимой, когда эта скорость низка.
Отсутствие температурных вариаций в мелких прудах, где температурные изменения носят неравномерный характер, Везенберг-Лунд (Wesenberg-Lund, 1926) тоже считал доводом в пользу своей теории.
Существенно заметить, что начало возникновения сезонных изменений формы как приспособлений к парению Везенберг-Лунд относит к послеледниковому периоду, считая, что наши теперешние пресноводные планктеры происходят от ледниковых форм, которые тогда жили в холодных водоемах с незначительной разницей температуры по сезонам.
«Все эти водоемы были населены одной и той же всюду доминирующей неизменно во все времена года расой, той, которая еще и теперь населяет местности, где еще господствует ледниковое время или которые лежат еще близко от границ льда. Когда позднее температура воды в водоемах поднялась, то параллельно с повышением температуры (и, следовательно, уменьшением вязкости воды) в течение длительного времени из года в год повышалась необходимость увеличить способность парения. Здесь-то и лежит начало сезонной вариации».
Вследствие повышения температуры и уменьшения грузоподъемности воды летом планктические организмы должны были тем или другим путем или приспособиться к требованию повышенной способности парения, или погибнуть. Так как стадии покоя затрудняли материнскому организму пребывание в пелагической области, то их образование прекратилось; это повело к изоляции колонии и к постоянству рас: результатом всего этого явилась ярко выраженная локальная вариация.
«Таким образом, мы можем как локальные вариации планктеров, так особенно их сезонные вариации рассматривать как своего рода феномен ледниковой эпохи. Этот своеобразный факт, что все наши летние расы зимой соединяются в одну и ту же расу, которая идентична той, которая держится круглый год в областях вокруг Полярного круга, является как бы пережитком отдаленных времен, сохранившимся в развитии наших теперешних рас.
Как хронологически сквозь меняющиеся времена, так и географически от полюса к экватору расположились планктические организмы в серии форм. Для кладоцер серии начинаются, с одной стороны (хронологически), короткими неуклюжими формами в ту эпоху жизни Земли, которая соответствует ледниковому периоду, с другой (географически), — в той области, которая лежит ближе к полюсу, однако эти формы кончаются в современности и в более теплых областях длинными узкими формами.
Первый шаг в этом ряду форм чисто биологической природы; он нашел свое морфологическое выражение только потом в строении тела организма. Предохраненные исключением оплодотворения от смешения, эти модификации должны были зафиксироваться, и результатом одинаковых и направленных в одну сторону воздействий внешних влияний было появление вышеупомянутых вертикальных и горизонтальных рядов форм; их отдельные члены в более старое время считались за самостоятельные виды или разновидности, в настоящее же время, как сказано выше, всех их соединяют в очень небольшое число видов» (Везенберг-Лунд, 1921: 24).
Изложенная выше теория парения Везенберг-Лунда и Оствальда и толкование с ее позиций сезонных вариаций развились в одно из центральных зданий классической планктологии.
В дальнейшем явление сезонных вариаций продолжало привлекать внимание исследователей и правильность этой теории была подтверждена на разных объектах (Ceratium, phyliopoda, Keratella).
Однако в последующем был обнаружен целый ряд фактов, противоречащих представлениям Везенберг-Лунда и Оствальда. Главные возражения против теории парения следующие. Еще в 1897 г. Лаутерборн в Рейне у Нейгофена находил весной 4-рогие, а летом — 3-рогие формы Ceratium hirundinella, факт, стоящий в противоречии с теорией парения, требущей как раз летом увеличения относительной поверхности тела, т. е. увеличения числа рогов. То же самое наблюдал Гюйер (Guyer, 1911) в швейцарских озерах.
Лаутерборн находил длиннорогие формы у Keratella cochlearis не летом, а зимой. На основании своих классических исследований над сезонными изменениями К. cochlearis Лаутерборн (Lauterborn, 1900, 1903) приходит к выводу, что температура имеет значение в процессе цикломорфоза постольку, поскольку она влияет на половой период (Sexualitat), причем именно половой период, т. е. внутренний фактор определяет цикломорфоз.
В том же духе высказывается Бенинг (1912) специально в отношении цикломорфоза у кладоцер, указывая, что «до сих пор половому периоду как причине изменений придавали слишком небольшое значение, тогда как для ясного понимания причин вариаций необходимо принимать во внимание не только внешние влияния, но и редуцирующее действие полового периода по крайней мере как адекватной причины вариаций».
Значение внутреннего фактора в том же смысле выдвигает также Гартман (Hartmann, 1918). В пользу такого взгляда свидетельствует то, что цикломорфоз наблюдается именно у тех организмов, у которых партеногенез чередуется с двуполым размножением (кладоцеры, коловратки). Сезонные изменения у форм, размножающихся только половым путем (копеподы), ничего общего с цикломорфозом не имеют.
У коловраток, например, из покоящегося яйца всегда выходит самка (и только самка), которая образует сравнительно крупные амиктические яйца, развивающиеся без оплодотворения. После ряда партеногенетических поколений, в течение которых от поколения к поколению происходят описанные выше цикломорфические изменения, образуются миктические яйца. Они отличаются меньшими размерами. Из неоплодотворенных миктических яиц вылупляются самцы; если миктическое яйцо оплодотворяется, то оно образует покоящееся яйцо, имеющее особое строение и обычно толстую оболочку. После периода покоя из покоящегося яйца выходит самка, и цикл повторяется. Амиктические яйца, таким образом, — облигатно-партеногенетические, миктические — только факультативно-партеногенетические.
Кречмар (Kratzschmar, 1908), исследовав ход цикломорфоза у К. quadrata в Верхнем озере, близ Лунца, установил, что он протекает несогласно с теорией парения. Весной из покоящихся яиц появляются формы с длинными задними шипами, соответствующие К. quadrata typica, далее у последующих генераций задние шипы все более и более уменьшаются и наконец исчезают, образуются последовательно формы, соответствующие К. quadrata brevispina, К. quadrata valga, К. quadrata curvicornis. Цикл заканчивается появлением самцов, половым процессом и образованием покоящихся яиц.
Рюе (Ruhe, 1912) в своей монографии, посвященной роду Bosmina, приводит ряд фактов, показывающих, что метаморфоз у босмин не подчиняется теории парения, так как при одном и том же изменении температуры различные босмины изменяются в совершенно противоположном направлении. Так, всем формам ряда coregoni (Ruhe, 1912) присуще общее летнее относительное удлинение передних антенн, а у форм ряда longispina, наоборот, характерным является летняя редукция этих антенн.
Таким образом, биологическое значение цикломорфоза босмин нельзя или по крайней мере не вполне можно раскрыть с помощью теории парения.
Точно также теории парения не подчиняется ряд фактов сезонных изменений у коловраток Keratella quadrata, K. cochlearis и polyarthra trigla, у ветвистоусых рачков Daphnia cucullata и D. pulex.
Существенным возражением против приписывания изменениям температуры воды решающей роли в возникновении сезонных вариаций служили эксперименты Вольтерека (Woltereck, 1909), показавшие, с одной стороны, возможность образования у дафний высокого шлема даже в холодной воде, но при достаточном снабжении их пищей, т. е. при усилении их интенсивности ассимиляции, с другой стороны, подавления развития шлема даже в теплой воде, но в условиях голодания.
Далее Вольтерек (Woltereck, 1913) указывает на то, что так называемые выросты парения, чтобы отвечать своему назначению, должны были бы располагаться перпендикулярно направлению погружения, тогда как, например, дафния при плавании ориентирует свою продольную ось, шлем и задний шип почти вертикально. Имеется немало кладоцер, говорит Вольтерек, у которых увеличивается не длинная ось тела, а перпендикулярно к ней стоящая спинно- брюшная ось, имеются также кладоцеры, которые одинаково удлиняются и в том и в другом направлениях.
Теория парения, на что обратил также внимание Вольтерек, не в состоянии объяснить самого возникновения так называемых органов парения, ибо последние выполняют свое назначение только тогда, когда они уже развиты сильно, в виде же зачатков они нейтральны и не имеют ценности для отбора.
И, наконец, Вольтерек (Woltereck, 1913) экспериментально показал, что «выросты парения» у дафний и босмин фактически представляют органы руления и стабилизации, обеспечивающие животному как активному пловцу возможность держаться в наиболее кормном слое. Так как глубокие озера летом обнаруживают резко выраженную слоистость водной толщи и в температурном, и в световом отношении, с концентрацией пищи в виде наннопланктона в верхнем слое, то кладоцеры как фильтраторы должны быть пространственно ограничены этим верхним наиболее кормным слоем. Этому сохранению уровня, сообщению путям плавания горизонтальности и прямолинейности и помогают имеющиеся у пловцов выросты как органы направления.
Теория плавания или руления.
Свои взгляды на значение организации и сезонных вариаций планктических ракообразных Вольтерек обосновал в своей чрезвычайно остроумной «теории руления», подвергнув детальному анализу функциональное значение выростов у босмины.
По Вольтереку (Woltereck, 1913), направление движения у кладоцер в воде является результатом, слагающимся из направления, сообщаемого ударом плавательных антенн (2-я пара антенн у кладоцер), и рулевого отклонения; к этому присоединяется еще влияние силы тяжести, действие света и механического раздражения от прикосновения к поверхности или ко дну. Сила тяжести оказывает погружающее действие на тело и изменяет положение длинной оси тела, подымая ее передний конец, если центр тяжести расположен кзади от середины тела. Всякое прикосновение и сотрясение тела кладоцер действует как раздражение на движение антенн. Свет также играет роль регулятора направления, воздействуя на глаз дафнии, который является своего рода фотостатическим аппаратом. Нарушение под влиянием светового раздражения оптического равновесия глаза вызывает сокращение глазных мышц, которое передается мышцам, управляющим движением антенн, последние, совершая определенное число взмахов, сообщают телу такое положение, при котором глаз пребывает в состоянии оптического равновесия. Различная интенсивность светового раздражения на глаз влияет на число ударов антенн у кладоцер: так, в глубокие сумерки Daphnia cucullata, по наблюдениям Вольтерека, производит 49 взмахов антенн в 1 минуту, при диффузном верхнем освещении — 72, при концентрированном верхнем освещении, в зависимости от расстояния от источника света — 88, 120, 138 взмахов. после трехминутного затемнения вслед за освещением — 56, новое освещение (диффузный верхний свет) — 80 взмахов в 1 минуту.
Следует при этом заметить, что взмах плавательных антенн у босмин направлен в сторону брюшка, тогда как у дафний он направлен в сторону спины. В результате босмина при каждом взмахе антенн свою голову поднимает, а дафния ее немного опускает.
Понять действие всех факторов, влияющих на направление плавания у кладоцер, лучше всего можно из анализа формы движения у босмины, как это проделал Вольтерек.
На рис. воспроизведено нормальное положение босмины при плавании.
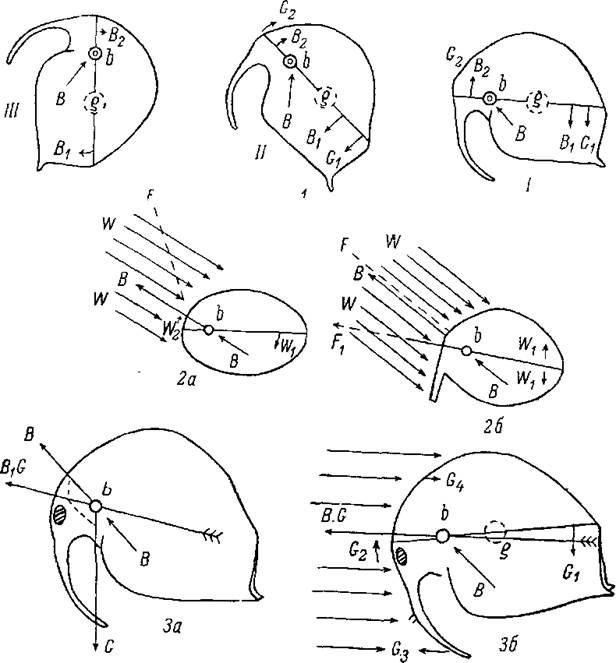
Продольная ось тела подпирается в точке b (основание антенн). В результате взмаха антенн в сторону брюшка происходит толчок в направлении В и поворот продольной оси в направлении В1В2. Одновременно действует сила тяжести, центр которой располагается на продольной оси несколько кзади от середины ее в точке g. Под воздействием этой силы произойдет поворот оси в направлении G1G2. В итоге действия двух сил — взмаха антенн и силы тяжести — босмина из положения i перейдет в положение ii. Действие тех же сил в новом положении тела должно вызвать положение iii, т. е. движение босмины будет происходить по окружности с перевертыванием через голову.
Так было бы, если бы такому движению не противодействовали бы некие силы, которые обеспечивают босмине горизонтальность пути ее нормального плавания.
Рассмотрим силу действия сопротивления (давления) воды на тело босмины и то, как она отражается на направлении движения босмины. Обратимся для этого к схеме (рис., 2а), на которой тело босмины изображено в виде простого овала без всяких выростов. Под влиянием взмаха антенн и силы тяжести произойдет передвижение тела в направлении В. Одновременно действует давление воды (W), распределяющееся неравномерно на верхнюю и нижнюю половины тела, именно, в данном случае, с перевесом давления на верхнюю половину (4 стрелки против двух). В результате такого распределения давления произойдет еще больший поворот продольной оси в направлении W1W2 с крутым подъемом ее переднего конца, и движение отклонится в направлении к F.
Прикрепив к передней части тела некоторую рулевую поверхность такой длины, чтобы давление, испытываемое телом со стороны воды, равномерно распределялось как на верхнюю, так и на нижнюю его половины, мы этим самым упраздним влияние этого фактора, и поступательное движение тела сохранится как направление пути, сообщенное действием взмаха антенн и силы тяжести (В=F).
Удлинив руль и направив его иначе, обусловив тем самым перевес давления воды на нижнюю половину тела, мы можем движению тела придать горизонтальное направление. Таким рулем у босмины и служит их хобот, образованный передней парой их антенн. Назначение хобота — противодействовать уклонению продольной оси от горизонтального положения ii поднятию головы, т. е. сообщение движению босмины горизонтального направления. Что, действительно, хобот босмии имеет такое назначение, доказывают последствия ампутации хобота: после такой операции движение босмины происходит по окружности с перевертыванием через голову.
Внимательное рассмотрение устройства тела различных видов Bosmina (В. longirostris, В. coregoni longispina, В. с. gibbera и др.) показывает, что длина и положение хобота у них всегда соответствуют длине и положению спины, благодаря чему соотношение между факторами, поднимающими голову (взмах антгин, сила тяжести, спинное сопротивление), и факторами, опускающими голову (давление воды), регулируется так, что движение становится горизонтальным.
В связи со сказанным наблюдается удлинение хобота при образовании летом горба на спине, укорачивание хобота зимой при отсутствии горба. При быстром движении (летом) для сохранения горизонтальности пути достаточно относительно короткого хобота; при замедленном движении (зимой) необходим более длинный хобот; если такого удлинения не происходит, движение совершается по кривой линии. Поэтому наблюдаются сезонные вариации в длине хобота; некоторые локальные формы В. lоngirostris и В. coregoni longispina летом имеют короткий, зимой — длинный хобот, факт, находящийся в противоречии с теорией парения, но вполне понятный с точки зрения теории руления. Летом эти животные плавают в более теплой воде и при обильной пище прямолинейно при помощи быстрых энергичных взмахов плавательных антенн; и холодной воде и при недостаточно интенсивной ассимиляции они передвигаются медленно, с половинной энергией, дугообразно, с частым поворотом на бок или на спину, и нуждаются поэтому в более длинной рулевой поверхности.
В других случаях видно, что — в интересах более быстрого поступательного движения — хобот, который, как всякий руль, сильно наклонен к направлению движения, одновременно играет роль тормоза, укорачивается и замещается мукронами, выростами, лежащими у заднего края обеих створок раковины и удлиняющимися большей частью параллельно направлению движения. Это имеет место у обычных летних вариаций В. longispina, а также В. reflexa.
В своей длине и положении мукроны также варьируют. У В. longirostris они направлены вниз, у В. coregoni (поскольку они имеются) — большей частью вытянуты назад. У первой они играют, как и хобот, роль руля, у второй выполняют функцию направительной плоскости, подобно брюшным краям створок раковины. В последнем случае вместе с краями раковины они затрудняют уклонение от направления пути и увеличивают вертикальную проекцию тела, в те же время оказывая некоторое противодействие рулевому отклоняющему действию хобота.
Интересно, что при укорачивании хобота наблюдается удлинение мукронов, которые в этом случае берут на себя функцию хобота. Сам хобот, когда он располагается в направлении движения, меняет спою функцию, становясь направительной плоскостью, особенно тогда, когда мукропы меняют стабилизирующую функцию на рулевую. Впрочем, обе функции часто переходят одна в другую. К ним присоединяется еще третья функция хобота— увеличение вертикальной проекции, но только у таких форм, которые свой хобот держат в значительной его части горизонтально.
Наглядным примером того, как меняются длина и положение хобота в связи с высотой спины у босмин, служат сезонные вариации Bosmina coregoni thersites. Зимой она имеет хобот нормального размера, чтобы тело ее при низкой спине могло перемещаться прямолинейно; летом xoбoт в соответствии с высокой спиной удлиняется, причем последняя возможно является приспособлением к парению, а вместо с удлиненным хоботом выполняет очень важную побочную роль, обеспечивая горизонтальность пути плавания.
Из произведенных анализов формы движения босмин и дафний с большой убедительностью вытекает, что своеобразие формы у планктических кладоцер вовсе не является приспособлением к парению и может рассматриваться как приспособление к плаванию. Главная функция выростов (шлем, хобот, горб и пр.) — регулирование направления движения при плавании, обсспсчиванис прямолинейности и горизонтальности пути, которое должно гарантировать животному прсбывание в наиболее кормном слое воды.
Кладоцеры в более глубоких водоемах с резко выраженной слоистостью должны быть пространственно ограничгны слоем эпилимниона, где имеется достаточное для них количество пищи (фитопланктона). Вольтерек считал, что в озере с температурной слоистостью оптимальные кормовые условия находятся в небольшом тонком слое, который лучше могут использовать только дафнии, способные к горизонтальному движению.
Так как дафнии при соприкосновении с зеркалом воды или при погружении в холодные слои гиполимниона погибают, то во избежание этого они должны иметь приспособления, препятствующие погружению и всплыванию. Назначение шлема и других выростов заключается в том, чтобы сделать передвижение более горизонтальным, удержать тело на определенном уровне. Следовательно, эти органы — плоскости направления, рули и стабилизаторы, причем выросты располагаются параллельно поступательному движению и при всяком уклонении действуют как противорули, затрудняя уклонение от прямолинейного и горизонтального направления пути.
Рассматриваемые морфологические приспособления, таким образом, у кладоцер служат отнюдь не для увеличения сопротивления формы. Например, у Bosmina coregoni var. gibbera спинной горб и увеличение летом поверхности раковины могут играть роль органа парения лишь в том случае, если рачок будет плавать на боку, т. е. наибольшей плоскостью тела перпендикулярно к направлению погружения. Однако рачок при плавании ориентируется как раз обратно — плоскость раковины (включая и спинной горб) располагается отвесно, параллельно направлению погружения, и, следовательно, никакого сопротивления при трении о частицы воды не оказывает. Это справедливо также для дафний и вообще для всех планктических кладоцер, имеющих сплющенную с боков раковину.
Вольтерек отмечает, что у планктических кладоцер повышение сопротивления формы обеспечивается только удлинением оси тела, в большинстве случаев увеличением морфологической продольной оси. Такую же роль выполняют у них распростертые горизонтально в фазе покоя антенны.
Парение большинства зоопланктеров в сущности является замедленным погружением. При этом первоначальное положение организма при плавании на определенном уровне восстанавливается за счет активных движений, регулируемых с помощью ряда приспособлений. У зоопланктона парение в смысле медленного погружения имеет место только тогда, когда выключается двигательный аппарат. Чтобы не опуститься совсем на дно, необходима работа двигательного аппарата, с помощью которого животное снова поднимается.
Такова сущность теории плавания, или руления, Вольтерека, как она изложена самим автором.
Теория «жизненности».
Теория «жизненности» разработана Ваглером (Wagler, 1923а, 19231), 1927, 1936) и занимает промежуточное положение между теориями парения и плавания. На основании своих наблюдений в природе и оценки зоогеографического распространения различных видов дафний Ваглер пришел к выводу, что колыбель всего рода Daphnia следует перенести в умеренную зону Евразии. Рассмотрев распространение каждого вида, наличие или отсутствие у них цикломорфоза в водоемах разного типа и на разных широтах, Ваглер установил следующее: цикломорфные дафнии встречаются только в крупных водоемах, но при любом климате. Так, на севере почти до Полярного круга находима D. cristata, в умеренной зоне — D. pulex, D. longispina, D. cucullata, D. barbata, D. lumholtzi и D. carinata, в тропиках — D. longispina, D. barbata, D. lumholtzi и D. carinata с их крайне выраженной изменчивостью. Уже такое географическое распространение цикломорфных дафний исключает признание за вязкостью воды роли единственного фактора, от которого зависят сезонные вариации.
Какова же истинная причина сезонных вариаций? Базируясь на наблюдениях над D. cucullata, а также на данных других авторов, Ваглер утверждает, что у D. cucullata мелкие расы свойственны мелким и плоским прудам, более крупные — средним прудам и небольшим озерам, а исполинские — преимущественно обитатели глубоких и крупных озер. Сходно ведут себя босмины и D. longispina. С крупными расами D. longispina всегда находятся в сообществе и крупные босмины (Bosmina coregoni) и крупные Daphnia l. galeata или D. l. pellucida. Средним расам D. cucullata соответствуют средней величины расы Bosmina coregoni (gibbera, thersites и др.), а В. соregoni никогда не встречаются в мелких прудах, в них встречаются лишь В. longispina и вместо D. galeata — очень мелкие нежные разновидности D. longispina.
Далее, оказалось, что у крупных D. cucullata больших озер половая зрелость достигается позже, чем у мелких прудовых форм — факт, известный и среди других животных, у которых значительные размеры тела и позднее наступление половозрелости связаны друг с другом.
Сопоставив размеры тела у D. cucullata с высотой головы, Ваглер пришел к заключению, что самые высокие головы имеют самые исполинские расы, среднего размера расы имеют и голову средней высоты, а круглоголовыми будут мелкие варианты этого вида.
Таким образом, не только признак наступления половозрелости и размеры тела связаны друг с другом, но сюда присоединяется еще и высота головы (шлема). А если вспомнить еще и то, что данный вид в своем географическом распространении не переступает некоторые (северную и южные) границы, т. е. связан с определенной средней температурой и, следовательно, с определенными условиями питания и дыхания, то это заставляет признать температуру со всеми вытекающими следствиями ответственной за образование изменчивых выростов тела.
Цикломорфоз должен обнаруживать наибольший размах там, где для вида имеются самые оптимальные условия существования в смысле тепла и пищи. Высокие шлемы у дафний являются, по Ваглеру, выражением наилучших оптимальных жизненных условий, процветания, благоденствия, жизненности вида в определенный отрезок времени в отдельном биотопе и даже во всей области обитания. Всякий переход за пределы этого оптимума в ту или другую сторону ведет к образованию не вариабильных (карликовых) рас (слишком сильно прогретых, сильно евтрофных мелких озер и прудов) или к полному исчезновению вида (в типично олиготрофных озерах Альпийской области и Арктики).
Ваглер обращает внимание на то обстоятельство, что «слой скачка в больших озерах с круто падающим береговым откосом лежит выше, чем в плоских водоемах с более широкой береговой отмелью; в первом типе поэтому эпилимнион менее глубок, чем во втором. Поскольку, как установлено, эпилимнион для тепловодной D. cucullata является местом ее обитания, то для нее жизненное пространство в озерах и прудах средней глубины мощнее, чем в крупных: опасность, что животное погрузится из прогретого эпилимниона в холодный гиполимпион в первых меньше, чем в последних. Поэтому в крупных, более холодных водоемах пути плавания для дафнии благодаря развитию очень высокого шлема становятся более горизонтальными, чем в озерах средней глубины и прудах.
В очень плоских водоемах вариации полностью исчезают не потому, что парение не дает никаких преимуществ, а потому, что вода равномерно прогрета, и опасность погружения в холодную воду исключена, максимум вариации иногда падает не на лето, а на весну или осень. Слой же скачка теперь мало- помалу расформировывается и медленно перемещается в глубину. Поэтому в одном и том же водоеме имеющееся в распоряжении дафнии пространство весной меньше, чем летом; точно так же начинающееся осенью охлаждение опять суживает зону обитания».
Так как в больших и глубоких водоемах жизненное пространство для пелагических кладоцер невелико, то не только с точки зрения теории жизненности, но и теории плавания приспособления для плавания и руления должны быть развиты больше, что и имеет место в природе.
Ваглер показал, что циклические изменения фирмы у D. cucallata связаны с циклами размножения; всегда весной первые генерации партеногенетического ряда украшены самыми высокими шлемами, позднее, несмотря на более высокую температуру, высота головы уменьшается. Это связано с тем, что весной слой скачка располагается близко к поверхности, слой эпилимниона небольшой толщины, и, следовательно, планктсры всего более нуждаются в хорошо развитых приспособлениях для руления. Осенью при падении температуры воды может снова иметь место увеличение шлема. Иногда летом может произойти несколько циклов вариаций.
Таким образом, Ваглер решающую роль в происхождении сезонных вариаций отводит температуре, но в ином смысле, чем Везенберг-Лунд и Оствальд. Главная причина, определяющая цикломорфоз, по Ваглеру, — ассимиляционные процессы, связанные с температурой и питанием, причем изменения вязкости воды с температурой сами по себе не представляют формообразующего фактора.
Результаты анализа изменений температуры воды и хода вариаций подтверждают взгляд Ваглера, заключающийся в том, что максимум вариаций имеет место только при оптимальной температуре, т. е. кривые хода температуры и вариации до тех пор синхронны, пока не перейден специфический для данной расы оптимум температуры: каждый подъем температуры обитаемого слоя до оптимума сопровождается пиком вариации, каждое опускание температуры ниже оптимума и поднятие выше оптимума, вызывая смещение среды в неблагоприятную сторону, вызывают уменьшение жизненности и редукцию органов парения, уменьшение размеров и количества яиц.
Смотря по температурной потребности данной расы, редукция изменчивых признаков будет наступать раньше или позже в году. Так, у многих холодноводно-стенотермических видов кульминационный пик вариации наступает в холодный сезон, а спад — уже ранней весной.
Разный ход вариации имеет место у частей популяции, приуроченных к различным глубинам с неодинаковой по высоте температурой одного и того же расчлененного озера.
Современная оценка теорий о причинах сезонных вариаций организмов планктона
При изучении живых организмов, подчиненных не одному частному влиянию, а воздействию большого числа внешних и внутренних факторов, нельзя какое-либо жизненное проявление приписывать влиянию только одного фактора.
Влияние такого рода будет определяться только при каких-то конкретных условиях, при других же условиях может четко проявиться действие другого фактора, маскируемое в данном случае. Поэтому при биологических исследованиях рискованно, выявив роль такого-то фактора при определенных условиях, обобщать его действие, забывая, что при изменившихся условиях может оказать действие совсем другой фактор, маскирующий влияние первого фактора.
Таким образом, указанные теории содержат только частицу истины, но, принимая во внимание сложность влияний, которым подвергаются организмы планктона, и разнообразие условий, в каких они живут, необходимо отметить у всех этих теорий один существенный недостаток, который заключается в том, что они выделяют один фактор и приписывают ему слишком широкое, чуть ли не универсальное значение. На самом же деле сезонные изменения организмов планктона обусловливаются не одним фактором, а их возникновение является результатом воздействия и плотности, и вязкости воды, и распределения пищи, и стабильности водной толщи, и освещенности и т. д.
По-видимому, у каждого отдельного биотипа образуется некоторая норма реакции, обнимающая сумму всех проявлений как внешних, так и внутренних воздействий, результатом которых у всех их, даже принадлежащих к разным родам, семействам и даже классам, получаются такие особенности строения, которые направлены к одной цели — к увеличению способности их к парению, причем различные приспособления к парению не только повышают способность собственно парения, по могут играть роль парусов или уловителей, перехватывающих токи в воде.
Сезонная изменчивость планктона при теперешнем состоянии исследования представляется как комплекс синхронно связанных изменений, которые в первую очередь вызываются температурой путем воздействия ее на жизненность планктических организмов. Что касается основного вопроса, именно более глубокого биологического значения цикломорфоза, то этот вопрос пока остается открытым.