Процесс пищеварения начинается в полости рта. И хотя продолжительность нахождения пищи в ротовой полости очень мала (в среднем 15-18 с), тем не менее, именно здесь осуществляются:
Ø измельчение пищи (благодаря разжевыванию). Процесс жевания запускается рефлекторно в ответ на раздражение рецепторов ротовой полости (тактильных, температурных, вкусовых) поступившим пищевым комком. Аффетрентная импульсация от рецепторов ротовой полости по чувствительным нервным волокнам лицевого (от передних 2/3 языка) и языкоглоточного (от задней трети языка) нервов достигает продолговатого мозга (солитарного ядра – общего чувствительного ядра для лицевого, языкоглоточного и блуждающего нервов). От солитарного ядра возбуждение передается к двигательному ядру тройничного нерва, заложенному в мосте и имеющему отношение к эфферентной иннервации жевательных мышц. Кроме того, в запуске жевания имеет немаловажное значение и некоторое растяжение жевательных мышц в связи с поступлением пищи в ротовую полость, которое вызывает рефлекторное их сокращение. Координацию жевания обеспечивают афферентные сигналы от проприорецепторов работающих жевательных мышц, сигнализирующие в ц.н.с. о силе и скорости мышечного сокращения.
Ø смачивание пищи слюной, что делает возможным начальный гидролиз пищевых веществ, а также облегчает последующее проглатывание. Слюна является пищеварительным соком, продуцируемым слюнными железами в полость рта. За сутки у человека образуется от 0,5 до 2 л слюны. Слюна вырабатывается тремя парами крупных слюнных желез (околоушными, поднижнечелюстными и подъязычными), расположенными вне слизистой ротовой полости, и множеством мелких слюнных желез, заложенных в толще слизистой языка, неба и щек. Выводные протоки всех слюнных желез открываются в полость рта. В зависимости от вырабатываемого секрета слюнные железы бывают трех типов:
ü серозные (вырабатывают жидкий секрет, не содержащий слизи – муцина) – околоушная железа и малые железы боковых поверхностей языка
ü слизистые (вырабатывают густую слюну, богатую муцином) – малые железы корня языка и неба
ü смешанные (вырабатывают серозно-слизистый секрет) – поднижнечелюстная и подъязычная железы, а также некоторые мелкие железы слизистой ротовой полости.
Слюна, находящаяся в ротовой полости, представляет собой результат смешения секрета всех слюнных желез и поэтому является смешанной. Вне приема пищи у человека слюна выделяется со скоростью 0,24 мл/мин для увлажнения слизистой ротовой полости, при жевании скорость слюноотделения составляет в среднем 3-3,5 мл/мин и зависит от вида принимаемой пищи. Около трети слюны, поступающей в ротовую полость за сутки, является продуктом секреции околоушных слюнных желез.
Слюна представляет собой вязкую мутноватую жидкость, состоящую из воды (99,4-99,5%) и сухого остатка (0,5-0,6%). Причем в сухом остатке слюны большую часть составляют органические вещества (их в 2-3 раза больше, чем минеральных солей). Органические вещества слюны отчасти являются продуктом обмена веществ самих секреторных клеток слюнных желез, а частично транспортируются из крови. В состав органических веществ слюны входят
· белки, в том числе ферменты – a-амилаза, некоторые протеиназы (катепсины, саливаин, гландулаин), липазы, кислая и щелочная фосфатазы, РНК-азы, фермент b-мурамидаза (лизоцим, обладающий бактерицидным действием), каллекреин (катализируют образование кининов – каллидина и брадикинина, которые обладают вазодилятаторным действием и оказывают местное сосудорасширяющее действие на артериолы слюнных желез, способствуя улучшению кровотока в них при повышении секреторной активности). Ферментативная активность пищеварительных ферментов слюны, кроме a-амилазы низкая, поэтому в ротовой полости начинается только гидролиз углеводов. В связи с очень кратковременным нахождением пищи в ротовой полости полисахариды под действием a-амилазы успевают расщепиться только до декстринов (полисахаров, содержащих до 10 моносахаридных остатков), при этом образуется небольшое количество и дисахарида мальтозы. Дисахаридазная активность слюны низкая, но все же при приеме дисахаридов они успевают в ротовой полости расщепиться до моносахаридов, после чего начинают всасываться в кровь.
· свободные аминокислоты и некоторые углеводы
· муцин – слизь, облегчающая проглатывание пищи
· ряд конечных продуктов метаболизма, подлежащих выделению из организма (мочевина, аммиак, креатинин и некоторые другие).
Реакция смешанной слюны (рН), как правило, находится в пределах 5,8-7,4 (слабо кислая – слабо щелочная) и во многом зависит от скорости секреции (при повышении скорости секреции рН увеличивается до 7,8). Ферментный состав, каталитическая активность, количество выделяемой слюны и ее вязкость подвержены значительным индивидуальным колебаниям и зависят от характера принимаемой пищи. Так, на сухую пищу выделяется более вязкая слюна и тем в больших количествах, чем суше принимаемая пища. На отвергаемые вещества, кислоты, горечи выделяется большое количество жидкой слюны. Такое четкое соответствие между характером и количеством принимаемой пищи и свойствами продуцируемой при этом слюны достигается благодаря нервным и гуморальным влияниям на слюнные железы. Причем в регуляции слюнных желез как начального отдела пищеварительного тракта наибольшее значение имеют нервные механизмы регуляции. Выделение слюны, как правило, начинается по типу условных рефлексов в ответ на раздражение зрительных, обонятельных или слуховых рецепторов (видом, запахом пищи или звуком тарелок и вилок). Эфферентная информация от соответствующих участков коркового представительства слюноотделительного рефлекса достигает пищевого центра гипоталамуса, а от гипоталамуса – направляется к парасимпатическим (ромбовидный мозг, где заложены верхнее и нижнее слюноотделительные ядра – парасимпатические ядра лицевого и языкоглоточного нервов) и симпатическим центрам (спинной мозг, боковые рога II-IV грудных сегментов) регуляции слюноотделения. При сильной возбудимости пищевого центра (что характерно для голодного человека) секреция слюны начинается через 1-3 с после раздражения рецепторов, тогда при слабом возбуждении пищевого центра – спустя 20-30 с. Возбуждение парасимптических нервов, иннервирующих слюнные железы, сопровождается образованием большого количества жидкой слюны, а длительное раздражение парсимпатических нервов приводит к истощению слюнных желез (при этом уменьшается содержание гранул в гландулоцитах слюнных желез, а в слюне – концентрация органических компонентов). Симпатический отдел и медиатор норадреналин стимулируют образование небольших количеств густой слюны, богатой органическими веществами (при этом усиливается образование и накопление гранул в гландулоцитах). Эффекты симпатического отдела вегетативной нервной системы возникают спустя более длительный латентный период, чем парасимпатические. Поступление пищи в ротовую полость и раздражение ею рецепторов ротовой полости (тактильных, вкусовых, температурных) вызывает дальнейшие слюноотделительные (но уже безусловные) рефлексы. Афферентная информация от рецепторов ротовой полости по чувствительным нервным волокнам лицевого и языкоглоточного нервов достигает солитарного ядра (чувствительного ядра) продолговатого мозга, а от этого ядра возбуждение передается к верхнему и нижнему слюноотделительным ядрам (вегетативным парасимпатическим ядрам лицевого и языкоглоточного нервов) и к симпатическим ядрам спинного мозга (II-IV грудных сегментов). Эферентная импульсация от парасимпатчисеких и симпатических центров слюноотделения оказывает стимулирующее влияние на секреторную активность, постоянно подстраивая количество и ферментный состав вырабатываемой слюны характеру принимаемой пищи.
Рефлекторные влияния на слюнные железы могут оказывать и тормозное влияние на их секрецию. Так, например, эмоции, умственное напряжение, дегидратация организма и некоторые другие факторы снижают активность пищевого центра гипоталамуса, тормозя тем самым и слюнную секрецию. Раздражение же пищевого центра высокой концентрацией угольной кислоты в крови (при гиперкапнии и асфиксии) сопровождается активацией пищевого центра и, как следствие, усилением слюноотделения
Ø анализ вкусовых свойств пищи (достигается благодаря наличию в слизистой ротовой полости – сосочках языка и других участках слизистой – вкусовых рецепторов, которые избирательно реагируют на определенные вкусовые качества). Такой анализ вкусовых свойств важен для нормального пищеварения (поскольку возбуждение вкусовых рецепторов рефлекторно стимулирует секреторную и моторную активность всего пищеварительного тракта), а также для формирования аппетита или возможного отказа от принимаемой пищи (при ее плохих вкусовых качествах)
Ø кроме того, раздражение пищевыми массами рецепторов ротовой полости (вкусовых, температурных, тактильных) играет важную роль для запуска секреторной активности не только слюнных желез, но и желез желудка, поджелудочной железы и печени, а также рефлекторно стимулирует моторику пищеварительного тракта, что необходимо для нормального переваривания принятой пищи.
После измельчения пищи в ротовой полости и смачивания ее слюной осуществляется проглатывание пищи, которое представляет собой рефлекторный акт, включающий три фазы:
Ø ротовую (быструю произвольную), в эту фазу благодаря сокращению мышц дна ротовой полости и языка из пищевого содержимого отделяется небольшой пищевой комок (объемом 5-15 см3), который в результате сокращения мышц языка и щек перемещается на спинку языка, а затем благодаря сокращению мышц языка переводится на корень языка (за передние дужки). Возбуждение рецепторов слизистой корня языка и неба рефлекторно вызывает сокращение
ü мышц, приподнимающих мягкое небо (что препятствует попаданию пищи в полость носа)
ü мышц самого языка, что обеспечивает определенные его движения, проталкивающие пищевой комок в полость глотки
ü мышц, смещающих подъязычную кость и вызывающих тем самым поднятие гортани (что препятствует попаданию пищи в дыхательные пути).
Сокращение мышц ротовой полости в момент ротовой фазы глотания приводит к повышению давления в ней, что облегчает переход пищевого комка в расслабленную глотку (давление в которой значительно ниже). Обратному возврату пищевого комка из глотки в ротовую полость препятствует сокращение мышц корня языка, приподнимающих язык и прижимающих его корень к передним дужкам
Ø глоточная (быстрая непроизвольная) фаза. Возникает рефлекторно в ответ на раздражение рецепторов глотки поступившим пищевым комком и проявляется в последовательном сокращении мышц глотки (в каждый данный момент сокращаются мышцы глотки, расположенные выше пищевого комка, что способствует локальному повышению давления в области пищевого комка и проталкиванию его вдоль глотки). В момент, когда пищевой комок достигает дистального отдела глотки, в результате локального сокращения мышц дистального отдела глотки и соответственно повышения давления в этом отделе, открывается глоточно-пищеводный сфинктер (благодаря тому, что давление в дистальном отделе глотки (45 мм.рт.ст) становиться выше такового в пищеводе (30 мм.рт.ст.)). Кроме того, именно потому, что давление в глотке выше давления в пищеводе, обратное забрасывание пищи из пищевода в глотку невозможно. Раздражение рецепторов ротовой полости и глотки в момент глотания рефлекторно вызывают не только сокращение их мышц, но и мышц, подтягивающих пищевод к зеву и расширяющих начальную его часть, куда и переходит пищевой комок из глотки. Ротовая и глоточная фазы как самые быстрые фазы глотания длятся всего около 1 с. Глоточная фаза является непроизвольной: ее нельзя выполнить, если в полости рта нет пищи или слюны
Ø пищеводная (медленная непроизвольная) фаза – самая длительная фаза глотания (составляет около 8-9 с для твердой пищи и 1-2 с для жидкой). Начинается с момента перехода пищевого комка в начальный отдел пищевода и соответственно раздражения рецепторов его слизистой оболочки, которое вызывает рефлекторное сокращение его мускулатуры (в начальной трети – скелетной, а в нижних 2/3 – гладкой). Причем сокращения мускулатуры пищевода носят характер перистальтической волны: последовательно сокращаются кольцевые мышцы стенки пищевода несколько выше места нахождения пищевого комка в направлении от проксимального отдела пищевода к дистальному его отделу. Такая перистальтическая волна способствует продвижению пищевого комка по пищеводу, а, дойдя до дистального его отдела, приводит к значительному локальному повышению давления в этой области, что способствует открытию пищеводно-кардиального сфинктера и последующему проталкиванию пищевого комка в кардиальную часть желудка. Кроме того, раздражение рецепторов слизистой дистальной части пищевода пищевым комком, рефлекторно вызывает не только сокращение гладкой мускулатуры пищевода в этой области, но и приводит к расслаблению мышц кардиальной части желудка, что способствует созданию довольно большого градиента давления между пищеводом и кардинальной частью желудка и открытию пищеводно-кардиального сфинктера (в отсутствии глотания это сфинктер закрыт). При наполнении желудка, наоборот, рефлекторно повышается тонус мышц кардии, что препятствует открытию пищеводно-кардиального сфинктера и соответственно забрасыванию пищи из желудка в пищевод.

Рис. 8. Схема акта глотания (обратите внимание, что во время глотания мягкое небо приподнято, что препятствует попаданию пищи в носовую полость, гортань приподнята, и вход в нее закрыт надгоратнником).
Центр глотания заложен в ромбовидном мозге и представлен
Ø солитарным ядром – общим чувствительным ядром для лицевого, языкоглоточного и блуждающего нервов. Это ядро получает чувствительную информацию от рецепторов слизистой оболочки рта, глотки, пищевода и многих других органов грудной и брюшной полости
Ø амбигуальным ядром – эфферентным соматическим ядром языкоглоточного и блуждающего нервов. Это ядро имеет отношение к иннервации скелетных мышцы языка, глотки и передней трети пищевода
Ø задним ядром блуждающего нерва – эфферентным парасимпатичеким ядром блуждающего нерва, обеспечивающим эфферентную иннервацию (наряду с прочими органами) гладкой мускулатуры пищевода.
Кроме того, в регуляции глотания могут участвовать и симпатические вегетативные ядра, заложенные в боковых рогах грудных сегментов спинного мозга и имеющие отношение к эфферентной иннервации гладкой мускулатуры пищевода и кардинальной части желудка. Так, если парасимпатические влияния стимулируют перистальтику пищевода и расслабляют кардию, способствуя пищеводной фазе глотания, то симпатические – оказывают противоположный эффект (тормозят моторику пищевода и повышают тонус кардии).
Пищеварение в желудке
Желудок представляет собой расширение пищеварительного тракта, расположенное в брюшной полости, интраперитонеально, между пищеводом и двенадцатиперстной кишкой. Причем желудок залегает в верхней части брюшной полости, под диафрагмой и печенью. Умеренно наполненный желудок имеет длину 24-26 см, а средняя его вместимость у взрослого человека составляет около 3 л.
В желудке выделяют:
Ø переднюю стенку, направленную кпереди (к передней стенки брюшной полости) и несколько кверху (в сторону диафрагмы)
Ø заднюю стенку, обращенную кзади (к задней стенки брюшной полости) и книзу (к области малого таза).
По краям желудка, где сходятся передняя и задняя его стенки, образуются:
Ø малая кривизна желудка (направлена вверх и вправо), в верхней ее части находится место впадения пищевода в желудок, называемое кардинальным отверстием.
Ø большая кривизна желудка (направлена вниз и влево).
В желудке выделяют следующие части:
Ø кардинальная часть желудка – область, прилежащая к кардинальному отверстию
Ø дно или свод желудка (фундальная его часть) – куполообразное выпячивание, расположенное слева от кардиальной части (обращено вверх (в сторону диафрагмы) и влево)
Ø пилорическая или привратниковая часть желудка – правый узкий отдел желудка, сообщающийся с двенадцатиперстной кишкой
Ø тело желудка – средняя часть желудка, расположенная между его кардинальной частью и дном слева и пилорической частью справа.

Рис. 9. Части желудка (вид спереди).
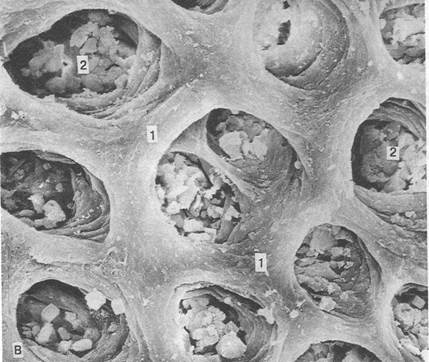
Рис. 10. Поверхность слизистой желудка (сканирующая электронная микроскопия)
1 – эпителий
2 – желудочные ямочки, на поверхность которых открываются протоки желудочных желез, вырабатывающих желудочный сок
Общий план гистологического строения стенки желудка аналогичен таковому всех остальных отделов пищеварительной трубки. Так, в составе стенки желудка выделяют три оболочки:
Ø наружную серозную, являющуюся производным висцерального листка брюшины и представляющую собой тонкий слой рыхлой волокнистой соединительной ткани с жировыми клетками, покрытый снаружи мезотелием
Ø среднюю мышечную, которая в желудке, в отличие от других отделов пищеварительной трубки, является не двух-, а трехслойной. Наружный слой мышечной оболочки образован продольными гладкомышечными волокнами, средний – круговыми, а внутренний – косыми
Ø внутреннюю слизистую оболочку с подлежащей подслизистой основой (прослойка рыхлой волокнистой соединительной ткани, имеющая довольно большую толщину). Слизистая оболочка желудка имеет некоторые особенности строения и включает следующие структуры:
ü однослойный однорядный кубический железистый эпителий, выстилающий желудок. Все клетки этого эпителия являются секреторными и продуцируют слизь (муцины) и бикарбонат-ионы (НСО3 --ионы). Причем секреция бикарбонат-ионов эпителиальными клетками на их поверхность осуществляется несколькими путями:
· выходом НСО3 – через апикальную мембрану клетки в обмен на Cl-, который транспортируется в клетку,
· активным транспортом НСО3 –, не зависящим от транспорта Cl-,
· пассивным транспортом НСО3 –через мембрану базальной части клетки внутрь клетки и по межклеточным щелям, зависящим от гидростатического давления в подэпителиальной соединительной ткани слизистой оболочки.
Таким образом, эпителий слизистой желудка, создавая слизисто-бикарбонатный барьер, обеспечивает защиту стенки желудка (в том числе и свою собственную защиту) от соляной кислоты, пепсинов и других потенциальных повреждающих агентов. Слой слизи имеет градиент рН: у его поверхности рН равен 2, а в примембранной части – 7. Кроме того, апикальная поверхность мембран эпителиальных клеток очень слабо проницаема для ионов Н+, а между латеральными поверхностями клеток имеется множество плотных контактов, создающих защитный барьер от проникновения чужеродных агентов (вирусов, бактерий), а также ионов водорода через эпителиальный пласт. Все это (наличие слизи на поверхности желудочного эпителия, градиент рН в слизи и плохая проницаемость апикальных мембран клеток для водорода) обеспечивают в норме хорошую защиту от повреждающего действия соляной кислоты. Но при некоторых условиях (механическом повреждении слизистой, воздействии на нее продуктов окисления, алкоголя, желчи) концентрация Н+ внутри клеток резко возрастает, что приводит к их гибели и нарушению слизисто-бикарбонатного барьера (разрушение барьера происходит в течение нескольких минут) и является одной из причин возможного дальнейшего изъязвления слизистой желудка. Повреждению барьера могут способствовать нестероидные противовоспалительные препараты (аспирин, индометацин, грамотрицательная бактерия Helicobacter pilori, выживающая в кислой среде желудка и поражающая поверхностный его эпителий). Для клеток железистого эпителия желудка характерна очень большая скорость физиологической регенерации (средняя продолжительность их жизни составляет около 3 дней)

Рис. 11. Механизм секреции бикарбонатов эпителиальными клетками слизистой желудка и двенадцатиперстной кишки.
А – выход НСО3- в обмен на Сl-
Б – активный транспорт НСО3-, не зависящий от транспорта Сl-
В и Г – транспорт НСО3- через мембрану базальной части клетки внутрь клетки и по межклеточным пространствам (зависит от гидростатического давления в подэпителиальной соединительной ткани слизистой оболочки)
ü собственную пластинку слизистой оболочки, образованную рыхлой волокнистой соединительной тканью с большим количеством лимфатических узелков. В этой пластинке слизистой заложены собственные железы желудка (всего их насчитывается около 35 млн.), продуцирующие желудочный сок, а также энтероэндокринные клетки, продуцирующие гормоны и гуморальные факторы желудка (гастрит, гистамин, внутренний фактор Касла и некоторые другие). Энтероэндокринные клетки могут располагаться как одиночно в собственной пластинке слизистой, так и входить в состав желудочных желез. Наибольшее количество желудочных желез заложено в области дна или свода желудка (т.е. в фундальной его части), а наибольшее количество энтероэндокринных клеток – в пилорической части.
ü мышечную пластинку слизистой, образованную тремя слоями гладкомышечных клеток.
Слизистая оболочка желудка и подслизистая основа формируют продольные складки, исчезающие в растянутом органе. Возможность образования таких складок обусловлена наличием довольно толстой подслизистой основы и мышечной пластинки в слизистой оболочке. В области перехода желудка в двенадцатиперстную кишку находится кольцевая складка – заслонка пилоруса, которая при сокращении пилорического сфинктера полностью закрывает выход из желудка. Вся поверхность слизистой оболочки желудка не является ровной: в ней имеется большое количество желудочных ямочек, представляющих собой устья многочисленных желез желудка (заложены в собственной пластинке слизистой).

Рис. 12. Гистоструктура стенки желудка
Гладкомышечные волокна желудочной стенки обуславливают его моторную функцию, которая обеспечивает
Ø временное депонирование пищи в желудке (резервуарная функция желудка)
Ø перемешивание слоя пищевых масс, прилежащего к слизистой желудка, с желудочным соком
Ø постепенное продвижение желудочного содержимого в направлении к привратниковой части желудка, откуда оно порциями эвакуируется в тонкий кишечник. Такое продвижение пищевых масс вдоль стенки желудка к выходу из него обеспечивается перистальтическими волнами сокращения, возникающими в кардинальной части (где находится кардиальный водитель ритма) и распространяющимися в направлении пилорической части. Благодаря перистальтическим волнам, распространяющимся в области тела желудка, тонкий слой пищевых масс, прилежащий к стенке желудка и обработанный желудочным соком, постепенно передвигается в направлении пилоруса.
Ø порционную эвакуацию пищевого содержимого, обработанного желудочным соком, в двенадцатиперстную кишку. Важную роль в такой порционной эвакуации играет моторика гладкомышечных волокон собственно привратниковой области желудка, благодаря которой создается высокое давление в области пилоруса, что способствует открытию привратникового сфинктера и переходу небольшой порции пищевого содержимого из желудка в кишечник. Большая же часть пищевого содержимого при этом в кишечник не переходит, но благодаря моторике пилорического отдела оно оказывается полностью перемешано с желудочным соком.
Железы желудка включают три типа клеток, отличающихся особенностями строения и продуцируемыми веществами:
Ø главные гландулоциты, синтезируют и секретируют пищеварительные ферменты – пепсиноген (предшественник пепсина) и липазу, имеют хорошо развитые гранулярную эндоплазматическую сеть и аппарат Гольджи, содержат большое количество зимогенных гранул в апикальной части (признак секреции клеткой белковых секретов)
Ø добавочные или слизистые гландулоциты, продуцируют слизь (муцин), в связи с чем характеризуются хорошо развитым аппаратом Гольджи
Ø обкладочные или париетальные гландулоциты, продуцируют соляную кислоту, содержат многочисленные митохондрии и систему внутриклеточных канальцев. Считается, что секреция париетальными клетками соляной кислоты на поверхность эпителия желудка является результатом работы нескольких ионных насосов (систем активного транспорта, транспортирующих ионы против концентрационных градиентов с затратой энергии АТФ). Так, через боковую поверхность клетки из межклеточных щелей в нее входит хлор в обмен на выводимый бикарбонат-ион (НСО3--ион). Такой обмен ионов обеспечивается работой Сl-/НСО3--насоса и, с одной стороны, делает возможным поступление в клетку хлора, а с другой – предотвращает изменение рН ее внутреннего содержимого (поскольку вход хлора сопряжен с одновременным выходом бикарбонат-иона). Поступивший в париетальный гландулоцит хлор затем выходит из него через апикальную мембрану клетки. В базальной мембране клетки встроен насос Na+ / Н+-АТФаза, транспортирующий внутрь клетки Н+ в обмен на выводимый натрий.В апикальной же мембране клетки имеется ионный насос – Н+/К+-АТФаза, выносящая ионы водорода из клетки в обмен на транспортируемые внутрь ионы калия. Регуляция внутриклеточной концентрации калия, который, с одной стороны, пассивно выходит из гландулоцита, а, с другой – поступает в клетку за счет работы Н+/К+-помпы, и натрия, пассивно входящего в гландулоцит и одновременно транспортируемого наружу с помощью Na+/Н+-помпы, достигается благодаря роботе Na+/К+-АТФазы. Таким образом, уже вне париетального гландулоцита секретируемые через ее апикальную поверхность ионы хлора и водорода ассоциируются друг с другом с образованием соляной кислоты. Причем работа двух главных насосов париетального гландулоцита – Н+/К+-АТФазы и Сl-/НСО3--насоса является тесно взаимосвязанной между собой. Так, при усилении работы Н+/К+-АТФазы и соответственно транспорта Н+ из клетки одновременно в 2-3 раза увеличивается и активность Сl-/НСО3--насоса, в результате чего внутриклеточный рН остается неизменным. Следует подчеркнуть, что в покое клетка секретирует только хлор, а Н+/К+-АТФаза является неактивной (т.е. соляная кислота за пределами клетки при этом не образуется). В железах желудка, расположенных в области пилорической (привратниковой) части, обкладочные гландулоциты отсутствуют, в связи с чем они не синтезируют соляную кислоту в составе желудочного сока, и сок привратниковой области имеет слабощелочную реакцию.
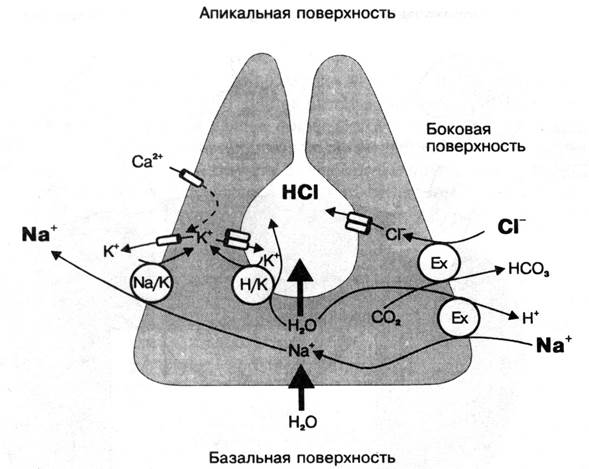
Рис. 13. Механизм транспорта ионов и секреции соляной кислоты париетальными клетками желудочных желез (Ех – обмен).

Рис. 14. Схема строения собственной железы желудка
Чистый желудочный сок, продуцируемый железами желудка, представляет собой бесцветную прозрачную жидкость с сильно кислой реакцией (рН=1,0-1,5), которая обусловлена наличием в нем соляной кислоты (ее концентрация в желудочном соке составляет 0,4-0,5%). При наличии пищи в желудке обычно концентрация соляной кислоты уменьшается (вследствие разведения пищевым содержимым, а также некоторой нейтрализацией НСl буферными системами принятой пищи), и соответственно рН желудочного сока увеличивается до 1,5-2,5 (а возможно и выше, что во многом зависит от буферных свойств пищи). Кислотность желудочного сока у ребенка первых месяцев жизни низкая (рН около 5,0), возрастает к концу года и достигает уровня взрослых к 7-12 годам. Соляная кислота желудочного сока обеспечивает:
Ø создание кислой среды, что необходимо для поддержания оптимальной активности протеолитических ферментов желудочного сока
Ø стимуляция секреции пепсиногенов главными гландулоцитами
Ø имеет отношение к активации пеписногенов (предшественники пепсинов – активных протеолитических ферментов). Наиболее вероятным считается предположение, согласно которому активация пепсиногенов происходит в присутствии соляной кислоты по механизму аутокаталитического действия самого пепсиногена, а именно путем отщепления от него короткого ингибиторного фрагмента, содержащего аргинин (в отсутствии же соляной кислоты пепсиноген свое аутокаталитическое действие не проявляет и остается неактивным ферментом)
Ø вызывает набухание белков (в результате чего увеличивается поверхность соприкосновения фермента с субстратом) и денатурацию белков, тем самым подготавливая их к ферментативному расщеплению
Ø обладает выраженным бактерицидным действием, выступая тем самым в роли одного из компонентов неспецифической защиты организма от чужеродных агентов (вирусов, бактерий), попадающих в организм с пищей.
Ферментативная активность желудочного сока обеспечивается следующими ферментами, синтезируемыми главными галндулоцитами желудочных желез:
Ø протеолитическими (являются эндопептидазами, катализирующими гидролиз лишь строго определенных пептидных связей в молекуле белка, поэтому под влиянием этих ферментов полного расщепления белков до аминокислот не происходит, а образуются лишь более короткие пептиды и небольшое количество свободных аминокислот). Все протеолитические ферменты вырабатываются в неактивном виде (в виде проферментов – пепсиногенов) и только в присутствии соляной кислоты желудочного сока активируются за счет проявления собственного аутокаталитического действия. В зависимости от того, при каком значении рН проявляют оптимум активности протезы желудочного сока, их делят на две группы:
ü пепсины (проявляют оптимум активности при рН 1,5-2,0, обладают выраженным протеолитическим действием в отношении многих белков и, в отличие от гастриксинов, обладают более выраженной протеолитической активностью в отношении белков молока)
ü гастриксины (проявляют оптимум активности в менее кислой среде – при рН 3,2-3,5).
Соотношение между пепсинами и гастриксинами в составе желудочного сока человека обычно составляет от 1:2 до 1:5. При рН свыше 5,0 протеазы желудочного сока полностью утрачивают свою активность.
ü ренин (содержится в желудочном соке детей грудного возраста, проявляет оптимум активности в слабо кислой среде (при рН 4,5), вызывает свертывание молока – превращение растворимого казеиногена в нерастворимый казеин, который в присутствии кальция выпадает в осадок, что приводит к резкому замедлению движения казеина по пищеварительному тракту и обеспечивает более длительное действие на него пртеиназ). Подобное свертывающее действие у взрослых людей выполняет сам пепсин.
Ø липазой, катализирующей расщепление преимущественно триглицеридов. В связи с тем, что липаза способна действовать только на эмульгированные жиры и при рН 5,5-7,5, ее роль в гидролизе жиров в желудке у взрослого человека не велика. Вместе с тем, у ребенка первого года жизни (рН желудочного сока составляет около 5,0), находящегося на грудном вскармливании и получающего уже эмульгированные жиры в составе грудного молока, липаза желудочного сока расщепляет до 25% жира молока. Кроме того, в расщеплении жиров грудного молока могут участвовать и липазы, входящие в его состав (аутолитическое пищеварение).
Кроме соляной кислоты и ферментов в состав желудочного сока входят
Ø мукоиды, продуцируемые добавочными гландулоцитами. Мукоиды желудочного сока, наряду с мукоидами, вырабатываемыми эпителиальной выстилкой желудка, обеспечивают механическую и химическую защиту его слизистой оболочки. К числу мукоидов, секретируемых в составе желудочного сока, относится и гастромукопротеид (внутренний фактор Касла), который необходим для всасывания в желудочно-кишечном тракте витамина В12 (цианкобаламина), который, в свою очередь, называют внешним фактором кроветворения, поскольку он поступает в организм извне (с пищей). Витамин В12 участвует в синтезе ДНК в момент усиленного деления клеток при кроветворении. Значение внутреннего фактора Касла для кроветворения состоит в следующем: он связывается с витамином В12 в желудке и защищает его от разрушения ферментами. Комплекс фактора Касла с витамином В12 в присутствии ионов Са2+ взаимодействует с рецепторами эпителиальных клеток дистального отдела подвздошной кишки. При этом витамин В12 поступает в клетку, а внутренний фактор высвобождается. Отсутствие фактора Касла приводит к развитию анемии. Большое значение для последующего транспорта витамина В12 кровью имеет транскобаламин II (транспортный белок, синтезируемый эпителием кишечника)
Ø неорганические вещества:
· минеральные слои (хлориды, сульфаты, фосфаты, бикарбонаты натрия, калия, кальция, магния)
· аммиак
Ø органические вещества:
· конечные продукты метаболизма (мочевина, мочевая кислота)
· аминокислоты, полипептиды.
Некоторые из этих веществ выделяются из слущихвшихся и разрушенных эпителиальных клеток слизистой желудка, а некоторые могут транспортироваться гландулоцитами из крови.
Желудочный сок, выделяемый разными участками слизистой оболочки желудка, имеет неодинаковую ферментативную активность и кислотность. Так, сок, вырабатываемый железами малой кривизны, отличается наибольшим содержанием пепсина и высокой кислотностью. Железы этого участка желудка первыми начинают секретировать сок, но и прекращают свою секрецию раньше, чем другие. Сок, вырабатываемый железами привратниковой части желудка, характеризуется небольшой ферментативной активностью, наличием большого количества слизи (в связи с чем представляет собой вязкую густую жидкость) и отсутствием соляной кислоты, в связи с чем имеет слабощелочную реакцию и не играет существенной роли в гидролизе питательных веществ, но обеспечивает частичную нейтрализацию кислой порции желудочного содержимого в пилорической части желудка перед эвакуацией ее в двенадцатиперстную кишку. Таким образом, желудочный сок является смесью соков, продуцируемых желудочными железами разных отделов