Однородность клеток в микробной популяции всегда относительна. Большую роль в процессах роста микробной популяции играет возрастная структура. Делиться, т.е. увеличивать численность популяции, способны только клетки, достигшие определенного возраста (или определенного размера). Возрастная гетерогенность популяции может служить причиной сложной немонотонной динамики ее численности.
Рассмотрим простейшую двухвозрастную модель клеточной популяции (Степанова, 1985). Популяция разбита на две группы клеток: молодые и старые.
Понятие “молодые” и “старые” применительно к разным видам микрорганизмов можно трактовать по-разному. В клетках эукариотов молодыми можно считать клетки G1, в которой синтезируется белок, а старыми ‑ все остальные, начиная с S -фазы синтеза ДНК. Именно на этих поздних стадиях существуют ингибирующие кейлоны, угнетающе действующие на скорость деления.
Будем считать, что клетки первой группы интенсивно растут, но не достигли физиологической зрелости и неспособны делиться. Члены второй группы способны к делению. Процесс деления может быть задержан при помощи различных ингибиторов. Уравнения для численностей молодых (N 1) и старых (N 2) клеток имеют вид:
 (11.27 )
(11.27 )
Здесь Т 1 - среднее время созревания молодой клетки,
Т 2 - среднее время пребывания старой клетки в репродуктивном периоде, D ‑ скорость протока. Удельная скорость деления клеток w=Т 2-1. Множитель 2 в первом уравнении отражает тот факт, что старая клетка делится на две молодые.
При отсутствии лимитирования субстратом продолжительность первой фазы Т 1 постоянна, продолжительность второй фазы Т 2 зависит от взаимного влияния клеток, которое осуществляется с помощью метаболитов (кейлонов), выделяемых клетками в среду. Если скорость выделения и распада кейлонов много больше скорости протока и скорости деления клеток, концентрация кейлонов пропорциональна числу клеток, их выделяющих.
Обозначим концентрацию ингибирующего метаболита I. Его влияние на удельную скорость деления клеток можно записать в виде:

Здесь n ‑ порядок ингибирования, k 1 -константа ингибирования.
Были рассмотрены три ситуации: 1) ингибиторы выделяются только молодыми клетками, 2) ингибиторы выделяются только старыми клетками, 3) независимо от возраста. Исследование модели показало, что только предположение о выделении ингибиторов старыми клетками позволяет описать колебательные режимы в системе. В рамках модели это означает, что скорость деления зависит от N 2:

Введем безразмерные переменные:

В безразмерных переменных система имеет вид:
 (11.28)
(11.28)
Штрих у времени опущен.
Кроме тривиальной особой точки система (11.28) имеет еще одну особую точку:
 ,
,
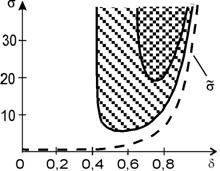
тип которой может быть различным в зависимости от параметров. Ширина области неустойчивости в пространстве параметров зависит от порядка ингибирования: чем больше n., тем она шире. Области неустойчивости на плоскости параметров (s,d) для второго и третьего порядка ингибирования изображены на рис. 11.5.
|
Фазовый портрет системы в области неустойчивости содержит предельный цикл (рис. 11.6).
|

Динамика переменных изображена на рис. 11.7.
|
