Значение отдаленной гибридизации в создании нового исходного материала особенно велико на современном этапе селекционной работы, когда мы имеем выдающиеся успехи в деле создания весьма ценных сортов различных сельскохозяйственных культур.
Для дальнейшего прогресса в выведении новых сортов, обладающих комплексом биологических, хозяйственно-полезных и корреляционно связанных с ними морфологических признаков, очень часто возникает острая необходимость выйти за пределы вида для заимствования необходимых свойств от других видов. Например, создание иммунных сортов картофеля к фитофторозу, вирусным заболеваниям, раку, нематоде, колорадскому жуку за счет внутривидовой гибридизации практически невозможно, так как все многообразие сортов и форм культурного вида Solanum tuberosum не обладают устойчивостью к этим заболеваниям и вредителям. Но среди других видов рода картофеля такие формы имеются. К ним, прежде всего, относятся S.demissum, S.acaule, S.andigenum, S.vernei, S.maglea и другие.
За прошедший период, особенно в XX веке, по теории и практике отдаленной гибридизации накоплен богатый экспериментальный материал, выяснены неиспользованные возможности ее более широкого применения, определены объективные трудности ее осуществления, выявлены основные причины существующих генетических и других биологических барьеров, разработано ряд методов преодоления нескрещиваемости видов между собой и бесплодия первого поколения отдаленных гибридов.
Большой вклад в теорию и практику этого вопроса внесли такие ученые как И.В. Мичурин (методы преодоления нескрещиваемости плодово-ягодных культур), Г.Д. Карпеченко (плодовитые капустно-редечные гибриды), Н.В. Цицин (сорта пшенично-пырейных гибридов), А.Ф. Шулындин (первые сорта ржано-пшеничных амфидиплоидов) и другие.
При изучении исторического становления и эволюции (филогении) различных видов растений установлено, что в основе линий многих видов лежит естественная, спонтанная гибридизация между видами. Наиболее богатым и убедительным примером на этот счет может служить эволюция всего полиплоидного ряда пшеницы (рис. 7) и создание человеком новой зерновой культуры тритикале с различными уровнями плоидности (рис.8).
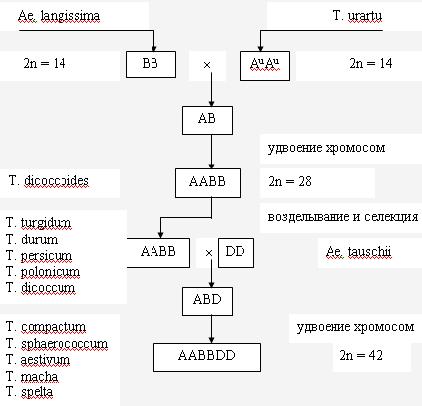
Рис. 7 Схема филогении рода Triticum L.
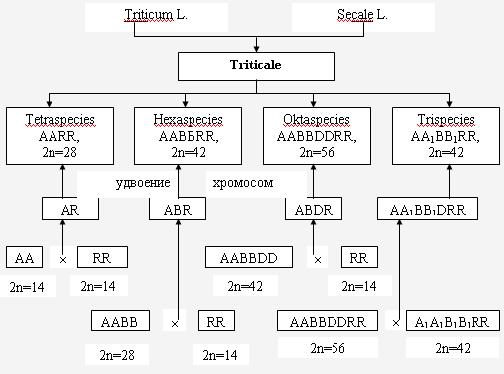
Рис. 8 Классификация и происхождение тритикале.
Эволюция видов пшениц группы эммеров шла по аналогичной схеме. Тетраплоидные эммеры T. araraticum и T. timopheevi (AbAbGG) произошли из гибридов T. boeticum (AbAb) и Al. speltoides (GG) через полиплоидизацию и отбор при размножении. Естественным гибридом и аллоплоидом от T. timopheevi и T. monocjccum является пшеница Жуковского (AuAuAbAbGG). Путем искусственной автополиплоидии французский ученый Эсло получил октоплоидную пшеницу T. timonovum (AbAbAbAbGGGG). Грибобойная пшеница T. fungcidum (AuAuAbAb BBGG) является результатом скрещивания T. persicum и T. ti
mopheevi, перевода гибридов на полиплоидную основу с отбором плодовитых потомств.
При проведении отдаленной гибридизации селекционеру постоянно приходится сталкиваться с тремя основными проблемами: нескрещиваемость генетически отдаленных видов, невсхожесть гибридных семян, бесплодие полученных гибридов.
Эти проблемы возникают в связи с тем, что:
- пыльца не прорастает на рыльце другого вида;
- пыльца прорастает, но пыльцевые трубки растут медленно и не достигают зародышевого мешка;
- пыльцевые трубки достигают зародышевого мешка, но оплодотворения не происходит;
- оплодотворение происходит, но зародыш прекращает свое развитие на стадии нескольких клеток;
- зародыш при начальном нормальном развитии прекращает свое формирование, образуются невсхожие семена;
- при физиологическом несоответствии цитоплазмы и чужеродных хромосом, отсутствии парности хромосом скрещиваемых видов коньюгация не происходит, мейоз нарушается, половые клетки не образуются и гибридные растения остаются бесплодными.
Заключение
В последнее время успешно решена проблема получения продовитого гибрида между пшеницей и ячменем, в результате чего получен новый вид зернового растения под названием Tritordeum.
Имеются примеры успешного применения отдаленной гибридизации в селекции табака, махорки и других культур.
Потенциальная возможность отдаленной гибридизации далеко еще не использована, предстоит раскрыть многие неизвестные механизмы взаимодействия цитоплазмы и ядра различных видов при их объединении в одну зачаточную клетку, предстоит более глубоко изучить физиологические и биохимические процессы, происходящие при прорастании пыльцы на рыльце чужеродного рыльца, продвижения пыльцевой трубки до семяпочки, слияния спермиев с центральным ядром и яйцеклеткой и всех процессов в постгамный период до полного формирования жизнеспособности семени.
Исключительно важную роль в дальнейшем совершенствовании и повышении эффективности отдаленной гибридизации должна сыграть биотехнология. С помощью ее методов культуры клеток и тканей возможна разработка способов извлечения из завязей оплодотворенных семяпочек и доращивания их на искусственном субстрате до получения гибридного растения, которое не может возникнуть обычным путем, так как во многих случаях через несколько дней после оплодотворения завязь с оплодотворенной семяпочкой отмирает. В последние годы доказана возможность получения отдаленных гибридов путем соматической гибридизации при слиянии клеток различных видов после их освобождения от клеточных оболочек с последующим получением каллуса, его дифференциацией до образования корней, листьев, стеблей и целых растений.
Список литературы
1. Основы генетики и селекции. / Под ред. Дубникова А.С. - М.: Колос, 1997.
2. Алтухов Ю. П. Генетические процессы в популяциях. - М.: Мир, 1983.
3. Замотайлов С. С., Бурдун А. М. Краткий курс генетики. - М.: Агропромиздат, 1987.
4. Дубинин Н. П. Общая генетика. - М.: Колос, 1987.
5. Биология. / Сост. Лемеза Н.А., Морозик М.С., Морозов Е.И. – Минск: Университетское, 1999. – с.-142.