С другой стороны, Великое Вымирание, возможно, было постепенным процессом, и, возможно, не имело никакого отношения к силам, приложенной внешними факторами.
Правда то, после мелового периода не существовало никаких динозавров, а другие существа, на которых воздействовало то, что тогда происходило и изменило окружающую среду, все исчезли в то же самое время. Однако, часто упускается из внимания тот факт, что эти животные и растения находились на пути к упадку задолго до этого. За одиннадцать миллионов лет до окончания мелового периода в Северной Америке было около трёх дюжин различных типов динозавров. К последнему миллиону лет этого периода то число сократилось вдвое. В одном известном североамериканском местонахождении динозавров слой, обогащённый иридием, был обнаружен, но последние кости динозавров лежат в породах значительно ниже его – представляя пробел от 20,000 до 80,000 лет – а развитие продвинутых палеоценовых млекопитающих началось в это время. Иридиевый слой, однако, кажется, совпадает с изменениями в растительной жизни. Донные морские отложения и породы из Техаса и Дании свидетельствуют, что иридий отлагался в течение периода продолжительностью до 100,000 лет – это слишком долго для любых отложений от метеоритного удара.

Влияние дрейфа континентов
На протяжении раннего мезозоя континенты располагались близко друг к другу, и значительные части их территории были покрыты мелководным морем. Тёплые поверхностные воды поддерживали изобилие растительной и животной жизни. В позднем мезозое континенты расходились и между ними образовывались глубоководные морские районы. Мелководные шельфовые моря сменялись более холодными глубокими морями, и вырастали горы. С исчезновением приповерхностных местообитаний морские растения и животные вымерли.
Также существует достоверное свидетельство в пользу того, что вымирания на суше происходили в иное время, нежели вымирания в океанах. Датировка событий такого плана по летописи окаменелостей известна своей трудностью. Один из методов, используемых в этом случае, состоит в изучении следов магнитного поля Земли в прошлом. Время от времени магнитное поле меняет полярность, север становятся югом и наоборот. Это оказывает воздействие на минералы, обладающие магнитными свойствами, которые образуются в горных породах того времени. Когда были изучены магнитные свойства минералов, составляющих горные породы, образовавшиеся на суше и в море в конце мелового периода, оказалось, что вымирание наземных животных произошло примерно через полмиллиона лет после вымирания морских животных.


Парниковый эффект
Различное соотношение газов, находящихся в атмосфере, может оказать значительное влияние на климат. В нормальных условиях температуры находятся в равновесии. Часть солнечного света, который достигает поверхности Земли, повторно излучается в космос в виде инфракрасного излучения. Увеличение доли двуокиси углерода или водяного пара в атмосфере приводит к тому, что значительная часть рассеиваемого инфракрасного излучения оказывается поглощённой и «пойманной» в атмосфере, и вследствие этого поверхность Земли нагревается.
Когда все эти точки зрения оказываются принятыми во внимание, наиболее вероятной причиной вымирания в меловой период мог бы стать непрерывно продолжающийся процесс движения литосферных плит. Поверхность Земли непрерывно движется. Кора и наиболее внешняя часть мантии – слоя, который образует значительную часть объёма Земли – непрерывно образуются и разрушаются. Новый материал формируется в океанах, вдоль океанических горных хребтов, в то время как старый материал разрушается, поглощаясь в океанических желобах. В то же самое время материки, находящиеся на этой поверхности, перемещаются в процессе её движения, сталкиваясь и раскалываясь.
На протяжении большей части Века Рептилий все материки были расположены близко к друг другу. Они начали раздвигаться в юрский период и были явно на пути к своему современному положению в меловой период. Это движение создало большие горные хребты глубоко в океанах, и вытесненная вода растеклась по краям материков в виде мелководных морей. Тёплый влажный климат, порождённый этими особенностями географии, был идеален для крупных рептилий того времени. Затем, в конце мелового периода, мелководные моря высохли и начали расти новые горные цепи, в частности, Скалистые горы.
Морские животные, которые эволюционировали, приспосабливаясь к жизни в тёплых мелководных морях, не могли выжить, поскольку их тепловодные местообитания были поглощены и смешались с холодными водами открытого океана. Микроскопические растения и животные с раковинами из карбоната кальция жили в приповерхностных водах. Теперь их численность снизилась, что соответствует внезапному исчезновению карбоната кальция в отложениях того времени. Меньшее количество растений в приповерхностных водах означало, что был теперь меньшее количество содержащейся в атмосфере двуокиси углерода превращалось в кислород. Более холодная вода также означает, что в море могло раствориться меньшее количество двуокиси углерода. Результатом этого было увеличение доли двуокиси углерода в атмосфере. Углекислый газ обладает свойством задерживать излучение тепла с Земли; результатом этого был парниковый эффект, и климатические условия стали намного теплее. Эти факторы могли бы объяснить соотношения изотопов, наблюдаемые в осадочных породах того времени.
Меньшее количество мелководных морей означает малое количество осаждаемых отложений, и, как следствие этого, неизвестны долгие последовательности осадочных пород, датируемые этим временем. Из-за этого можно было бы считать продолжительность некоторого геологического отрезка времени значительно меньшей, чем она была фактически. Точно так же весь иридий, который был в то время извергнут из недр Земли вулканами, особенно вдоль линии молодых Скалистых гор, мог сконцентрироваться в том небольшом количестве отложений, которые тогда сформировались, что дало бы в результате аномально высокую концентрацию.
На суше площади материков стали больше, потому что заливавшие их мелководные моря высохли. Области суши, которые какое-то время были разделены, теперь были связаны сушей. Например, животные могли расселяться по Северной Америке в долготном направлении, хотя ранее они были разделены мелководным центральным морем. Они могли мигрировать по сухопутному мосту, который теперь появился между Азией и Северной Америкой. Поскольку животные из различных природных зон теперь смешивались друг с другом, они конкурировали за те же самые пищевые ресурсы. Они также принесли с собой болезни и паразитов, к которым у них был иммунитет, но у их новых соседей не было. В свою очередь, они также получили болезни и паразитов от своих соседей. Растительная жизнь также изменялась из-за меняющихся климатических условий.
В целом это было трудное время для любого живого существа. Если бы случилось так, что метеорит или комета столкнулись бы с Землей в это время, то это бы нанесло завершающий удар по уже разваливающейся системе.
Альтернатива...
Ради целей этой книги ничего из этого не случилось. Метеорит промахнулся. Облако комет прошло мимо. Морские растения смогли приспособиться к изменяющимся температурам воды. Наземные животные оказались способными сопротивляться болезням и паразитам своих соседей.
Короче говоря, животная жизнь, которая развивалась на протяжении 150 миллионов лет мезозойской эры, продолжила развиваться в течение, по крайней мере, ещё 65 миллионов лет без перерыва. И это стало основанием для нашей книги.
Однако, прежде, чем мы взглянем на тех созданий, которые существуют в нашей альтернативной зоологии современности, мы должны сначала посмотреть на то, каким животным был динозавр, и как он эволюционировал и развивался в эпоху своего расцвета, в отдалённые времена мезозойской эры.
ЧТО ТАКОЕ ДИНОЗАВР?
ЭВОЛЮЦИЯ НАЗЕМНЫХ РЕПТИЛИЙ
Согласно учебнику, понятие «динозавр» относится к любому представителю отрядов Saurischia и Ornithischia, двум из пяти отрядов в составе надотряда рептилий Archosauria.
Более доступным для понимания описанием было бы следующее: динозавр – это кто-то из числа обычно крупных наземных рептилий, которые были очень обычны на протяжении мезозойской эры; эта эра геологического времени включает триасовый, юрский и меловой периоды, между 284 и 65 миллионами лет назад. Ключевым словом в описании является понятие «наземный». Морские рептилии того времени – длинношеие плезиозавры, похожие на китов плиозавры, дельфинообразные ихтиозавры, морские ящерицы мозазавры, а также все черепахи – не были динозаврами. Ими не были ни летающие рептилии – птерозавры, ни пресноводные жители – крокодилы, хотя они представляют собой два близкородственных отряда в пределах надотряда Archosauria. Мезозойская эра была, несомненно, Веком Рептилий.
| * Последние находки говорят, что зверообразные рептилии продержались гораздо дольше триаса. Некоторые их группы были обычны в юрский период. А из отложений палеоцена Северной Америки известен Chronoperates – существо примитивного строения, которого некоторые палеонтологи считают зверообразной рептилией. Если это так, то оказывается, что зверообразные рептилии пережили динозавров. Есть даже мнение, что современные утконос и ехидна – это не млекопитающие, а зверообразные рептилии. – В. П. |
Архозавры (представители надотряда Archosauria) появились в процессе эволюции в конце пермского периоде. Это было время, когда зверообразные рептилии были главными наземными животными. Зверообразные рептилии были группой рептилий, совершенно отличающихся от архозавров, которые из маленьких ящерицеобразных животных развились к триасовому периоду в покрытых шерстью собакоподобных существ. В конце этого периода они вымерли*, оставив своих крошечных потомков, млекопитающих.
В течение всего времени, пока зверообразные рептилии были самыми распространёнными среди наземных животных, архозавры, которые также эволюционировали от маленьких ящерицеобразных существ, не были важной группой. Первым появившимся отрядом архозавров были Thecodontia (см. страницы 12-15). Это были довольно похожие на крокодилов существа, и многие вели образ жизни, сходный с крокодильим, в ручьях и реках. Только когда зверообразные рептилии вымерли, архозавры смогли распространиться и развиться в четыре отряда*.
| * Д. Диксон несколько упрощает ситуацию, существовавшую в триасе. Вымирание крупных зверообразных рептилий началось уже в раннем триасе, а в среднем триасе уже появились первые крокодилы, динозавры и птерозавры. В то же время ещё продолжали существовать разнообразные зверообразные рептилии – растительноядные дицинодонты и много групп плотоядных тероморфов разного размера. Иными словами, развитие архозавров началось на фоне достаточного разнообразия зверообразных рептилий. – В. П. ** Среди водных архозавров, кроме крокодилов, существовала своеобразная группа фитозавров. Они были хищниками и внешне отличались от крокодилов положением ноздрей – они открывались не на конце морды, а вблизи глаз. – В. П. |
У обитающих в воде текодонтов развились сильные задние лапы, приспособленные для плавания, и длинный гребущий хвост**. Когда эволюционировали наземные формы, они обладали способностью ходить на сильных задних ногах, а тело уравновешивалось длинным хвостом. Они продолжили вести жизнь хищников. Они образовали второй отряд архозавров - Saurischia или ящеротазовые динозавры. Ящеротазовые не все были двуногими плотоядными, а некоторые стали растительноядными. Поскольку растительноядность требует большего объёма кишечника, чем плотоядность, тело этих животных стало крупнее, и они больше не могли балансировать на своих задних ногах. Эти ящеротазовые приобрели четвероногое положение тела и у них развились длинные шеи.
Другая группа, которая произошла от текодонтов, способных ходить на задних ногах. Они были растительноядными, но могли сохранять равновесие на двух ногах из-за устройства своих бедренных костей. Объёмистый кишечник мог находиться скорее ниже бёдер, нежели перед ними. Это были Ornithischia – птицетазовые динозавры. С течением времени многие птицетазовые также утратили двуногое положение тела и стали четвероногими. Для них также отмечается тенденция к развитию разных видов панцирей впечатляющего облика.
Тем временем другая группа потомков текодонтов приспособилась к жизни в небе. Они начали как планирующие формы, но скоро превратились в летающих существ очень изощрённой формы, с крыльями, которыми могли взмахивать, маленьким негибким телом, стройными полыми костями и сложной нервной системой. Они были представителями отряда Pterosauria.
И последняя группа архозавров осталась очень консервативной, вела тот же самый образ жизни, что и их текодонтные предки – грелись на берегах рек и питались рыбой. Их форма тела очень мало изменилась. Они стали Crocodylia, единственной группой архозавров, которая пережила позднемеловое вымирание (согласно традиционной палеонтологии).

Эволюция динозавров
В позднем палеозое эволюционировало много различных групп рептилий. Наиболее успешными среди них были зверообразные рептилии. Как только они вымерли, за исключением линии, которая привела к появлению млекопитающих, архозавры распространились и стали самой важной группой.

Классификация динозавров
Динозавры классифицируются на основании строения их бедра. Ящеротазовые динозавры названы так из-за строения тазовых костей, где подвздошная кость прикрепляется к позвоночнику, лобковая кость направлена вперёд, а седалищная кость направлена назад, как у ящерицы. Плотоядные ящеротазовые ходили на двух ногах. Растительноядные ящеротазовые с тяжёлым брюхом ходили на четырёх ногах. Птицетазовые динозавры обладали лобковой костью, обращённой назад напротив седалищной кости, подобно тому, как это имеет место у птиц. Птицетазовые все были растительноядными с брюхом, свисавшим ниже бёдер. Многие панцирные формы обладали сильно разросшейся подвздошной костью для поддерживания веса панциря.
Динозавры правили Землёй на протяжении 170 миллионов лет, они доминировали над всеми другими формами жизни, и они были наиболее успешной из когда-либо существовавших групп живых существ. Что сделало их столь могущественными, столь доминирующими, столь успешными? В основном они были очень напористой и постоянно эволюционирующей группой. Как только природные условия менялись на протяжении мезозойской эры, они менялись, чтобы приспособиться к ним. Где были пустыни, были динозавры, живущие в пустыне. Где были болота, были динозавры, живущие в болотах. Леса породили динозавров, живущих в лесах. Вероятно, также были динозавры, живущие в горах, хотя это трудно установить, поскольку горные животные редко сохраняются в ископаемом состоянии.
Есть высокая вероятность того, что многие динозавры были эндотермными; у них был метаболизм, который мог позволить им регулировать свою температуру тела независимо от внешней среды. Это – состояние, которое обычно обозначается как «теплокровность» и имеется у млекопитающих и птиц. Это даёт животному очень высокую эффективность, и, хотя теплокровное животное нуждается в большем количестве пищи, эта пища может быть быстрее преобразована в энергию, которая может использоваться в течение более долгих периодов времени. Быстрые, активные, питающиеся мясом ящеротазовые динозавры весьма вероятно были теплокровными, поскольку только теплокровное животное могло быть достаточно проворным и активным, чтобы вести образ жизни этих существ. Большинство теплокровных животных покрыто мехом или перьями, чтобы обеспечить надёжную терморегуляцию. Птерозавры, крылатые родственники динозавров, определённо были теплокровными, и, насколько известно, покрытыми шерстью. Возможно также, что теплокровные динозавры могли обладать мехом или же пухом*. Сложно сказать, однако, были ли крупные растительноядные динозавры теплокровными. Аргументом против этого является то, что форма головы и шеи сделала бы невозможным для растительноядного существа с длинной шеей поедать достаточно пищи, чтобы обеспечивать потребности его тела при теплокровном образе жизни. Огромная масса тела, вероятно, в любом случае могла бы сохранять сравнительно постоянную температуру без всякой сложной системы поддержания теплокровности. Однако, поперечные срезы через ископаемые кости крупных растительноядных динозавров дают основание полагать, что они были быстрорастущими животными – и эта черта имеется только у теплокровных существ.
| * Во время написания этой книги (1988 год) не было прямых свидетельств, указывающих на строение покровов тела динозавров. Но в последнее время появились находки, свидетельствующие о наличии у динозавров перьев (Beipiaosaurus, Caudipteryx, Sinosauropteryx). Перья динозавров по строению сильно отличаются от перьев птиц. В книге Д. Диксона, однако, динозавры покрыты не перьями, а шерстью. Выходит, здесь он не угадал. – В. П. |
В плане анатомии динозавры имели очень совершенное строение. У других рептилии ноги выглядят торчащими по бокам, а тело подвешено между ними. Это достаточно эффективно для маленького животного наподобие ящерицы, но не достаточно для поддержки массивной туши динозавра. У динозавров в процессе эволюции развилось вертикальное положение, похожее на то, что есть у современного млекопитающего. Другими словами, ноги располагались вертикально под телом, и вес тела приходился на верхние части ног.
Скелеты самых крупных динозавров были шедеврами инженерных решений природы. Ноги были массивными, чтобы выдерживать большой вес, но позвоночники состояли из лёгких полых распорок, расположенных так, чтобы максимальная прочность обеспечивалась минимальной массой.
В нашей альтернативной зоологии Великое Вымирание не происходило. Поэтому все эти тенденции продолжили развитие. Динозавры эволюционировали и приспособились к меняющимся условиям Земли. Динозавры, а также другие крупные рептилии в воздухе и в море, всё ещё являются наиболее успешными и широко распространенными животными на поверхности Земли. Теперь, прежде, чем мы исследуем мир крупных рептилий, который мог бы быть сегодня, давайте посмотрим на историю животных начиная с конца мелового периода 65 миллионов лет назад. Давайте проследим эволюционное развитие гигантских рептилий в течение последующих третичного и четвертичного периодов.
НОВОЕ
ДРЕВО ЖИЗНИ
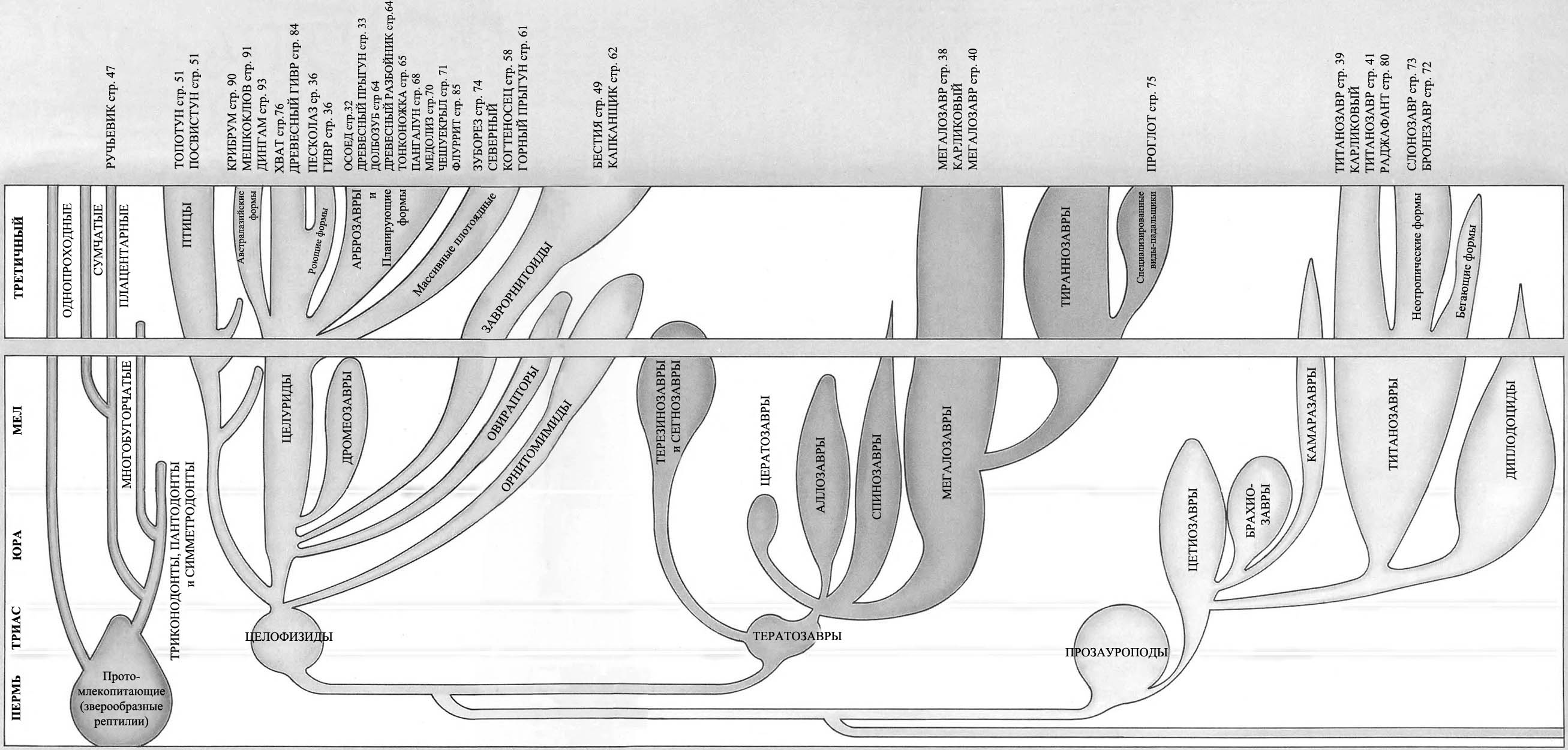
Продолжение родословной линии динозавров на протяжении последних 65 миллионов лет подразумевало, что эти великолепные существа процветали, расселялись и увеличивали своё разнообразие с той же самой энергией как и на протяжении предыдущих 150 миллионов лет. Более того, фактически, на протяжении последних 65 миллионов лет они были свидетелями стремительных изменений на поверхности Земли.
Большинство родословных ветвей динозавров фактически вымерло намного раньше конца мелового периода, и динозавры, которые живут сегодня, происходят от наиболее успешных из позднемеловых групп.
Легко сложенные плотоядные ящеротазовые, или «целурозавры», произошедшие от целофизид, процветали, будучи очень универсальными и способными к большому количеству эволюционных изменений.
Плотоядные тяжёлого телосложения, или «карнозавры», произошедшие от тератозавров, жили не столь хорошо. К концу мелового периода процветали только огромные тираннозавры. Мегалозавры сохраняют свою численность, но линия карнозавров не показывает значительного прироста.
Прозауроподы вымерли в конце триаса, но их потомки – тяжёлые длинношеие растительноядные ящеротазовые, или «зауроподы», стали обычными в юрский период, но в позднем мелу существовали только в местах, откуда они не были вытеснены орнитоподами. Те, что выжили до сегодняшнего дня, представляют собой главным образом потомков титанозавров и большей частью живут на южных материках.
Мелкие двуногие птицетазовые – «орнитоподы» – которые эволюционировали от гетеродонтозавров (страницы 14 – 15), подобно мелким плотоядным ящеротазовым, адаптировались и процветали, с лёгкостью приспосабливаясь к значительным изменениям окружающей среды. Большинство травоядных сегодняшнего дня эволюционировало из их числа.
Цератопсы – рогатые птицетазовые – были очень успешными в конце мелового периода и всё ещё встречаются в тех или иных формах сегодня.
Стегозавры – птицетазовые, имеющие костные пластины – почти все вымерли к меловому периоду, за исключением изолированной группы в Индии. Группа не пережила ледниковый период.
Анкилозавры – покрытые тяжёлой бронёй птицетазовые – были успешной группой мелового периода и выжили.
Ещё есть летающие птерозавры. Несмотря на конкуренцию с птицами, птерозавры выжили.
Среди впечатляющих водоплавающих рептилий длинно- и короткошеие плезиозавры, а также мозазавры выжили, но рыбообразные ихтиозавры вымерли.
А как же млекопитающие? Они начинали в триасовый период как маленькие насекомоядные существа. К концу мелового периода они всё ещё были маленькими насекомоядными существами. Они не имели возможностей расселяться и увеличивать разнообразие, и являются маленькими насекомоядными существами и по сей день*.
| * Дугал Диксон сильно обижает млекопитающих своими словами. К концу мелового периода млекопитающие были очень разнообразными, и он не мог этого не знать. Среди них существовали однопроходные, сумчатые, насекомоядные, кондилартры (предки копытных). Возможно, в это же время появились грызуны. Также существовало несколько архаичных отрядов млекопитающих, в том числе многобугорчатые, дожившие в нашем мире до эоцена. Самые крупные млекопитающие мелового периода, возможно, достигали размеров волка. Экология мезозойских млекопитающих была разнообразнее той, что приписывает им автор книги – среди них существовали не только насекомоядные формы, но и хищники и растительноядные. – В. П. |
| ||||
 Скелет целофизиса Coelophysis Целофизиды были основной триасовой группой плотоядных ящеротазовых динозавров лёгкого телосложения. От них произошли очень похожие и успешные целуриды и птицы, птицеподобные орнитомимиды и питающиеся яйцами овирапториды.
Скелет целофизиса Coelophysis Целофизиды были основной триасовой группой плотоядных ящеротазовых динозавров лёгкого телосложения. От них произошли очень похожие и успешные целуриды и птицы, птицеподобные орнитомимиды и питающиеся яйцами овирапториды.
|  Скелет дейнониха Deinonychus Дромеозавры и заврорнитоиды были двумя более специализированными ветвями целофизид. Обе группы включали свирепых охотников, для каждой из них характерен убивающий добычу коготь на задней лапе. Заврорнитоиды были особенно успешными.
Скелет дейнониха Deinonychus Дромеозавры и заврорнитоиды были двумя более специализированными ветвями целофизид. Обе группы включали свирепых охотников, для каждой из них характерен убивающий добычу коготь на задней лапе. Заврорнитоиды были особенно успешными.
|  Скелет мегалозавра Мегалозавры были в числе самых ранних, но наиболее успешных тяжело сложенных плотоядных. По сложению они напоминали группы плотоядных, существовавшие менее продолжительное время, вроде огромных аллозавров, спинозавров, имевших парус на спине, и рогатых цератозавров.
Скелет мегалозавра Мегалозавры были в числе самых ранних, но наиболее успешных тяжело сложенных плотоядных. По сложению они напоминали группы плотоядных, существовавшие менее продолжительное время, вроде огромных аллозавров, спинозавров, имевших парус на спине, и рогатых цератозавров.
|  Скелет тираннозавра Tyrannosaurus Тираннозавры был в числе последних эволюционировавших массивных плотоядных, и не появлялись до мелового периода. Они развились из линии мегалозавров и превратились в самых больших и самых тяжёлых плотоядных животных из когда-либо существовавших.
Скелет тираннозавра Tyrannosaurus Тираннозавры был в числе последних эволюционировавших массивных плотоядных, и не появлялись до мелового периода. Они развились из линии мегалозавров и превратились в самых больших и самых тяжёлых плотоядных животных из когда-либо существовавших.
|  Скелет апатозавра Apatosaurus Существовало несколько семейств длинношеих травоядных ящеротазовых динозавров. Самыми ранними были цетиозавры, брахиозавры и диплодоциды. Камаразавры существовали дольше, но также были титанозавры более лёгкого телосложения, которые в конечном счёте оказались наиболее успешными.
Скелет апатозавра Apatosaurus Существовало несколько семейств длинношеих травоядных ящеротазовых динозавров. Самыми ранними были цетиозавры, брахиозавры и диплодоциды. Камаразавры существовали дольше, но также были титанозавры более лёгкого телосложения, которые в конечном счёте оказались наиболее успешными.
|

| |||||||
 Скелет гетеродонтозавра Heterodontosaurus Гетеродонтозавры были исходной группой двуногих растительноядных птицетазовых динозавров, появившейся в триасе. От них эволюционировало несколько групп, которые были успешными и существовали дольше.
Скелет гетеродонтозавра Heterodontosaurus Гетеродонтозавры были исходной группой двуногих растительноядных птицетазовых динозавров, появившейся в триасе. От них эволюционировало несколько групп, которые были успешными и существовали дольше.
|  Скелет гипсилофодона Hypsilophodon Гипсилофодонты были, возможно, наиболее успешными из мелких двуногих травоядных. Они эволюционировали в юрский период как бегающие животные лёгкого сложения, быстроногие и способные быстро убежать от врагов.
Скелет гипсилофодона Hypsilophodon Гипсилофодонты были, возможно, наиболее успешными из мелких двуногих травоядных. Они эволюционировали в юрский период как бегающие животные лёгкого сложения, быстроногие и способные быстро убежать от врагов.
|  Скелет паразауролофа Parasaurolophus Появившись в позднемеловое время, гадрозавры стали наиболее распространёнными крупными двуногими травоядными. Они рано разделились на обладавших гребнями ламбеозаврин и не имевших гребня гадрозаврин.
Скелет паразауролофа Parasaurolophus Появившись в позднемеловое время, гадрозавры стали наиболее распространёнными крупными двуногими травоядными. Они рано разделились на обладавших гребнями ламбеозаврин и не имевших гребня гадрозаврин.
|  Скелет стегоцераса Stegoceras Пахицефалозавры появились и распространились в позднем меловом периоде. У них были массивные черепа, и они напоминали других двуногих травоядных, за исключением тяжёлых костных образований на их головах.
Скелет стегоцераса Stegoceras Пахицефалозавры появились и распространились в позднем меловом периоде. У них были массивные черепа, и они напоминали других двуногих травоядных, за исключением тяжёлых костных образований на их головах.
|  Скелет трицератопса Triceratops Цератопсы были рогатыми динозаврами – успешной группой, которая эволюционировала в меловой период. Они произошли от пситтакозавров – семейства двуногих травоядных, похожих на гипсилофодонтов.
Скелет трицератопса Triceratops Цератопсы были рогатыми динозаврами – успешной группой, которая эволюционировала в меловой период. Они произошли от пситтакозавров – семейства двуногих травоядных, похожих на гипсилофодонтов.
|  Скелет стегозавра Stegosaurus Динозавры, покрытые панцирем, разделяются на две главных группы – покрытых пластинами стегозавров, которые были наиболее успешными в юрский период, и покрытых шипами нодозавров, а также анкилозавров, которые широко распространились в меловой период.
Скелет стегозавра Stegosaurus Динозавры, покрытые панцирем, разделяются на две главных группы – покрытых пластинами стегозавров, которые были наиболее успешными в юрский период, и покрытых шипами нодозавров, а также анкилозавров, которые широко распространились в меловой период.
|  Скелет птеранодона Pteranodon Птерозавры, как и динозавры, произошли от текодонтов. Они начали летать в триасовый период. Ранние длиннохвостые рамфоринхоиды вымерли в юрский период и был замещены короткохвостыми птеродактилями.
Скелет птеранодона Pteranodon Птерозавры, как и динозавры, произошли от текодонтов. Они начали летать в триасовый период. Ранние длиннохвостые рамфоринхоиды вымерли в юрский период и был замещены короткохвостыми птеродактилями.
|  Скелет плезиозавра Plesiosaurus Плезиозавры с ластообразными конечностями были наиболее успешными из морских рептилий, существовавшими более долгое время, чем рыбообразные ихтиозавры. Было две главных линии эволюции плезиозавров – длинношеие эласмозавры и короткошеие плиозавры.
Скелет плезиозавра Plesiosaurus Плезиозавры с ластообразными конечностями были наиболее успешными из морских рептилий, существовавшими более долгое время, чем рыбообразные ихтиозавры. Было две главных линии эволюции плезиозавров – длинношеие эласмозавры и короткошеие плиозавры.
|
КРАКЕН (страница 107) и КОКОСОВЫЙ ВОР (страница 99) – беспозвоночные, которые происходят от головоногих моллюсков, потому они не включены в эту схему.
ПАЛЕОГЕОГРАФИЯ
ВЕЧНО МЕНЯЮЩИЙСЯ ЛАНДШАФТ
На протяжении основной части Века Рептилий материки раздвигались. В течение триасового периода все материки Земли были соединены вместе в один огромный суперконтинент под названием Пангея, состоявший из северной части, Лавразии, и южной части, Гондваны. В течение юрского периода этот суперконтинент начал раскалываться, и в меловом периоде разделяющиеся материки отчётливо находились на пути к их современному положениям.
На протяжении последних 65 миллионов лет, начиная с конца мелового периода, самое большое движение материков происходило в южном полушарии. В самом конце мелового периода Австралийский материк всё ещё соединялся с Антарктическим. Впоследствии он откололся и переместился в северном направлении, пока не достиг своего нынешнего положения в южном тропическом климатическом поясе. Континентальный блок, который теперь включает Индию, путешествовал в северном направлении через Индийский океан, и в конце концов столкнулся с огромным северным континентом. Поскольку он откололся от своего первоначального положения как часть восточноафриканского массива суши, он отделил также другие материковые фрагменты, которые остались после этого как острова, известные как Мадагаскар и Сейшельские острова, а также множество рассеянных погружённых фрагментов. Море, называемое Тетис, всё ещё отделяло материки, которые мы теперь называем Европой и Азией, от Африки, но это море медленно закрывалось и сужалось. На другой стороне земного шара два американских материка были довольно изолированы, и связывались только цепочкой островов на протяжении самого раннего третичного времени, и постоянным сухопутным перешейком, появившимся совсем недавно. Северная Америка, однако, почти постоянно соединялась с Азией через Берингийский сухопутный мост – перешеек, который только недавно погрузился в океан.
Движение материков оказало влияние на климат. Когда существовал непрерывный морской пояс вокруг Земли, состоящий из Тетиса и пролива между Америками, существовало постоянно текущее на запад экваториальное течение, которое образовывали господствующие ветры. Оно формировало тёплые и влажные климатические условия на окраинах большинства материков. Вода, нагретая в этом течении, циркулировала в океане, доходя вдоль береговых линий до севера северного полушария и далеко вниз до Антарктиды на юге. Климатические условия были тёплые и ровные, и влажные леса росли на большинстве материков.
Из-за движения материков море Тетис закрылось. В это же самое время Австралазия отошла далеко от Антарктиды и открылся циркумполярный морской путь, позволяя непрерывному течению, направленному на восток, окружить холодный Антарктический материк. Из-за всего этого пропало экваториальное течение, а вместе с ним и тёплые климатические условия. Теперь тёплые тропические воды меньше смешивались с холодными полярными, и климатические условия всё больше и больше различались на обширных территориях. Более сухие и прохладные условия, ставшие результатом этих событий, означали, что пышные тропические леса начали уступать место травянистым равнинам – новой среде обитания.