Согласование инноваций и адаптаций при периодической колонизации городской среды: молекулярные, генетические базисы и основы развития
Alexander V. Badyaev
Перевод: Шопина Ольга, студентка 1 курса магистратуры кафедры геохимии ландшафтов и географии почв географического факультета МГУ
Введение
Эволюция инноваций требует не только включения новых генетических или экологических факторов, но и поддержания функциональности уже созданных структур. Какова структура систем организма, которая обеспечивает такое сосуществование? Как новые и новые модификации могут распространяться и развиваться, прежде чем приобретут полную функциональность? Как новые признаки появляются во время развития? Ответы на эти вопросы требуют прямого анализа развития во время возникновения новых адаптаций, во время окна, когда полезный новый признак может быть произведен с помощью систем развития и генетических систем, но выживание организма все еще зависит от существующих адаптаций.
Здесь я обращаюсь к этим вопросам, изучая морфологическую эволюцию инвазивного вида, который в ходе колонизации новых сред постоянно развивает адаптацию к городской среде. В то время как колонизация различных сред у этого вида была связана с эволюцией новых адаптаций к новым источникам пищи, периодическая колонизация городской среды привела к конвергентной эволюции морфологии, наиболее полезной в городских условиях. Меня особенно интересовало взаимодействие между историческим непредвиденными обстоятельствами и текущим выбором развития адаптивных морфологий в городских условиях.
Из нескольких калифорнийских птиц, выпущенных из зоомагазина в Нью-Йорке в 1939 году, и в ходе естественного расширения его западного ареала, всего за несколько десятилетий мексиканская чечевица (Carpodacus mexicanus) стала занимать одну из самых широких экологических ниш среди всех существующих птиц (Badyaev et al., 2012). Первоначально птица жарких пустынь и сухих открытых мест на юго-западе, теперь встречается почти во всех типах ландшафтов и климатов в Северной Америке, от крайней северной тайги до океанских побережий и столичных районов. Однако часто упускается из виду тот факт, что во время колонизации их чрезвычайно разнообразного экологического ареала, мексиканские чечевицы также неоднократно колонизировали, из разных исходных точек, более однородную городскую среду.
Здесь я специально сосредотачиваюсь на быстрой эволюции морфологии клюва. У птиц, питающихся семенами, адаптация клюва отражает не только конкурентные требования обработки и дробления семян (Boag & Grant, 1981; Bowman, 1961; Willson, 1971), но также влияет на конфигурацию и вокализацию голосовых путей (Podos et al., 2004; Slabbekoorn & Smith, 2002). Например, большие клювы менее подходят для быстрой модуляции звука, такой как трели, тогда как более длинные клювы и более широкий зазор ограничивают эффективную длину голосового тракта и соответствующие частоты песен (Nowicki, 1987).
Существуют две причины, по которым колонизация городской среды особенно сильно влияет на морфологию клюва. Во-первых, чечевицы в недавно сформировавшейся городской популяции сильно зависят от кормушек; все расширение ареала до городских районов, особенно в крайних северных частях ареала (например, на юго-западе Аляски), стало возможным благодаря ассоциации с кормушками (Badyaev et al., 2012). Во-вторых, в кормушки обычно насыпают семена (такие как семена подсолнечника или крупного проса), которые намного крупнее и тверже, чем те, которые составляют основную часть рациона мексиканской чечевицы в естественных популяциях (Badyaev et al., 2008; Hensley, 1954). Такой контраст особенно очевиден при сравнении городского и сельского населения в ареале предков вида на юго-западе США: мелкие семена трав и кактусов, составляющие основную часть рациона семян в пустыне, имеют средний диаметр в направлении растрескивания 1,2 ± 0,4 мм (n = 377 семян) и средняя твердость 2,7 ± 0,08 Ньютона (Н), тогда как подсолнечники, которые являются основной частью рациона семян в городских районах, имеют средний диаметр в направлении растрескивания 6,8 ± 2,1 мм (n = 120) и среднюю твердость 8,11 ± 0,21 Н (Badyaev et al., 2008; Mills et al., 1989).
В этой главе я сначала опишу морфологическую дивергенцию и естественный отбор конфигураций клюва в городских и естественных популяциях. Во-вторых, я рассмотрю способы, с помощью которых инновации и адаптации могут быть согласованы во время адаптивной диверсификации клювов. В-третьих, я расскажу о развитии и генетической основе таких циклов дивергенции и конвергенции с городскими адаптациями и путях, с помощью которых онтогенетические механизмы могут выполнять точные, разнообразные и все же обратимые адаптации в конфигурациях клюва. Я предполагаю, что как городским адаптациям, так и дивергенции популяции способствовала модульная организация развития клюва, где небольшие регуляторные изменения в консервативных факторах молекулярного роста и значительная функциональная избыточность в полученных конфигурациях приводят к широкому диапазону адаптивных модификаций клюва. В заключение я проведу предварительный анализ молекулярной основы такой адаптивной эволюции и направления будущих исследований.
Периодические городские адаптации во время расширения экологической ниши
Быстрое и обратимое развитие локальных адаптаций, которое сопровождало расширение экологической ниши мексиканской чечевицы в Северной Америке в течение последних 70 лет, предоставляет прекрасную возможность непосредственно исследовать природу изменений в развитии во время критической, но обычно подробно не расматриваемой стадии эволюции - когда первоначальная фенотипическая адаптивность превращается в локальную адаптацию и когда существующие адаптации должны сосуществовать с новыми модификациями. В северных, недавно освоенных частях ареала популяции мексиканской чечевицы дают возможность изучать повторяющуюся эволюцию адаптаций к городской среде из разных начальных точек из-за различным непредвиденных исторических обстоятельств (рис. 1.1), а также в то время, когда необходимость быстрой реорганизации сложных клювов совпадает с необходимостью поддерживать некоторую стабильность оптимальной конфигурации клювов. В южных, унаследованных частях ареала дифференциация мексиканской чечевицы между городской и сельской средой дает нам представление о долгосрочных последствиях адаптации к городской среде в морфологических, культурных аспектах и системах развития (Badyaev et al., 2008). Оба временных отрезка будут исследованы здесь.
12.2.1 Различные экологические начальные точки для развития городских адаптаций
У мексиканских чечевиц пути колонизации новых сред не соответствуют местам наилучшей пригодности среды обитания для конкретных видов (рис. 1.2). Чтобы установить это, мы создали подробные географические информационные системы (ГИС) с разрешением ячейки = 1 км для > 300 местообитаний в недавно сформированных популяциях на северо-западе Соединенных Штатов, с использованием подхода на основе взвешенных расстояний, реализованным в пакете ESRI ARC/INFO spatial analysis (Badyaev and Krebs, неопубликованные данные). Вкратце, мы включили показатели видоспецифической устойчивости к движению и рассеиванию путем взвешивания расстояния между источником и целевой ячейкой на основе значений сопротивления промежуточных ячеек (Koen et al., 2012; Nikolakaki, 2004) и специально сфокусировались на параметрах которые описывают антропогенное развитие и типы растительности, потому что оба определяют основные изменения в рационе семян мексиканской чечевицы (например, однородные и крупные семена подсолнечника от кормушек по сравнению с мелкими и разнообразными местными семенами в сельской местности). Мы рассчитали экологическое сходство популяций на основе слоев ГИС дорог и жилищ, плотности населения (перепись Соединенных Штатов 2000 г.), высоты местности почв и типов растительности (50 типов местообитаний, разрешение выходных ячеек = 90 м), чтобы установить, что популяции в условиях последовательной колонизации (например, рисунок 1.1) испытывает исключительно разнообразные экологические условия всего за несколько поколений (рисунок 1.2). Значимость этого открытия заключается в том, что колонизация городских районов в результате таких вторжений происходит из сильно отличающихся экологических исходных точек.

Рис. 1.1 Пример последовательности городских и сельских популяций, сформированной в результате «скачков расселения» после формирования исходной популяции (например, сельское популяция CRW сформировалась из городской популяции MSO за 12 поколений) в северо-западной части Соединенных Штатов. Ландшафтные данные и ГИС (сопротивление промежуточных ячеек по данным разных ландшафтных слоёв) получены по (Badyaev & Krebs, не опубликованые данные). Климатические данные взяты за 1948–2010 гг. Морфометрические данные взяты из (Badyaev, 2010).
12.2.2 Быстрая и обратимая эволюция локально-адаптивных морфологий клюва.
Колонизация экологически различных областей во время расширения ареала была связана с быстрой реорганизацией морфологий клюва как внутри, так и между новыми популяциями (Badyaev & Hill, 2000; Badyaev & Martin, 2000). В конечном итоге эти изменения были вызваны тесной ковариацией состава рациона семян и силы укуса, что, в свою очередь, тесно связано с развитием костей и мышц клюва у этого вида (Бадяев, 2010, 2011а).
Долгосрочное исследование эволюции клюва в городских условиях (Badyaev, 2010) выявило три основных принципа. Во-первых, эволюция точных локальных конфигураций клюва не уменьшила будущие модификации; отличные и локально стабильные конфигурации оставались обратимыми, часто через много поколений в будущем. Во-вторых, функциональная эквивалентность различных конфигураций клюва обеспечила начальное выживание и позволила широко исследовать морфопространство клюва до того, как наиболее подходящая конфигурация была найдена и стабилизирована естественным отбором. В-третьих, в течение первых нескольких поколений в новом экологическом регионе чечевицы продемонстрировали быструю реорганизацию и разнообразие в конфигурациях клюва с последующей стабилизацией конфигурации клюва, соответствующей местным условиям (рис. 1.3).
Важно отметить, что стабилизация и начало местной адаптации по времени совпали с повторяющимися закономерностями расселения на краю географического ареала чечевицы, так группы взрослых особей проводят «скачки расселения» в новом месте после размножения, часто за сотни километров и осваиваются там и обитают круглогодично – это основной механизм расширения ареала у этого вида (Badyaev et al., 2012). В таких рассеивающихся стаях, которые в основном состоят из пар взрослого населения, не наблюдается постоянной дискриминации одного пола (Badyaev et al., 2001); эти «скачки расселения» происходят каждые 5–10 поколений после формирования популяции. Когда такие «скачки расселения» совпадают с возникновением и стабилизацией локально-адаптивной конфигурации клюва в исходных популяциях, они позволяют непосредственно изучить влияние начальной конфигурации на последующую эволюцию (рис. 12.3).
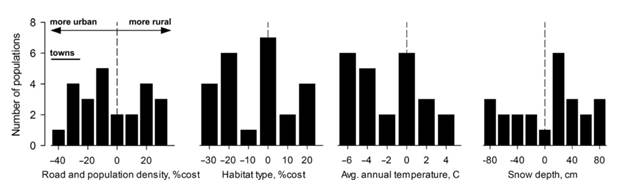
Рисунок 1.2 Разнообразные экологические условия, которые формировали городские адаптации в 24 недавно появившихся популяциях мексиканской чечевицы на северо-западе США. Показаны различия от исходных популяций (пунктирные линии) в плотности человеческого населения (как параметр для определения количества кормушек), по типу среды обитания (% разницы в пригодности типа среды обитания для популяции, полученные из слоев ГИС,% вес), по среднегодовой температуре и высоте снежного покрова (Badyaev & Krebs, неопубликованные данные).
12.2.3 Развивающая и функциональная избыточность конфигураций клюва
В целом, большая сила укуса (обусловленная, главным образом, шириной клюва) и более длинные клювы были более предпочтительны в городских районах, где рацион мексиканской чечевицы состоит почти исключительно из семян подсолнечника (Badyaev et al., 2008). Однако, несмотря на сильное изменение в силе прикуса как в сельских, так и в городских адаптациях, не было единой оптимальной конфигурации клюва, которая была бы предпочтительна в каждой среде (рис. 1.3c). В городских популяциях вклад физической формы (пропорционального отношения длины и ширины клюва при колеблющемся направленном отборе) различался между поколениями, в то время как глубина клюва имела характер стабилизирующего отбора (рис. 1.3).
Три вывода иллюстрируют адаптивную эквивалентность различных конфигураций клюва во время эволюции и преобладание компенсаторных взаимодействий развития и функциональности между компонентами клюва (Badyaev, 2010). Во-первых, несмотря на значительные микроэволюционные изменения в процессе адаптации к городской среде, морфологическая интеграция - мера развития и функциональной корреляции между компонентами клюва - оставалась сходной. Во-вторых, несмотря на широко различающиеся фенотипы клюва между поколениями во время адаптивных последовательностей (например, рисунок 1.1), изменение общей ковариационной структуры между поколениями было постепенным между соседними поколениями, ожидаемый паттерн, связывал последовательное микроэволюционное изменение в одном компоненте клюва (например, глубина клюва) с компенсаторным ростом в других компонентах. Наконец, детальный анализ фенотипической ковариационной структуры клювов во время эволюции выявил ее равномерность (Badyaev, 2010), подразумевая, что разные компоненты клюва демонстрируют компенсаторную вариацию в разных поколениях.
Основное эволюционное значение компенсаторных взаимодействий развития и функциональной универсальности в конфигурациях компонентов клюва заключается в том, что они поддерживают существенную вариабельность развития и генетические различия в отдельных компонентах, тем самым обеспечивая быстрые микроэволюционные изменения в процессе формирования популяции. Такая организация может не только обеспечить быструю сборку подходящей на местном уровне морфологии для эксплуатации ресурсов в новых средах, но также может согласовать необходимость специализации и тесной функциональной интеграции в клювах с обширной эволюционной диверсификацией после изменений в естественном отборе.

Рисунок 1.3 Морфология клювов чечевиц в городских популяциях является более однородной, независимо от популяции предков и местных климатических условий, чем морфология клювов чечевиц в коренной / сельской популяции. (a) Дивергенция между городскими (белые столбцы, n = 1024) и сельскими/ аборигенными (черные столбцы, n = 221) популяциями в юго-восточной Аризоне. (b) Недавнее расхождение между городскими популяциями (белые столбцы, n = 8740, пять групп популяций) и сельскими популяциями (черные столбцы, n = 9130, 11 групп популяций) на северо-западе Монтаны. (c) Поверхность соответствия (выживаемость по силе укуса) популяции МSО в течение 20 лет адаптации к городской среде, определяемая осями m1 (глубина клюва) и m3 (относительная длина и ширина) (Badyaev, 2010). Линии соединяют последующие поколения. Пунктирные эллипсоиды показывают две отличные области высокой пригодности - «городской оптимум» - в этой городской популяции. «Прыжjr расселения» и основание сельской популяции CRW совпадает с 12-м поколением эволюции адаптаций у городской популяции МSО.
12.2.4 Последствия для пения
Комбинация местной адаптации к поиску пищи, воздействия на вокальный аппарат и культурное наследование песен может сильно усилить локальную адаптацию в конфигурации клюва (Grant & Grant, 1996; M acDougallShackleton & MacDougall-Shackleton, 2001; Mayr, 1939). Для мексиканских чечевиц такое усиление может быть особенно эффективным по трем причинам. Во-первых, черты клюва, на которые больше всего влияют модификации в городских условиях - длина клюва и общий размер, - наиболее важны для создания звуков жужжания и трели - элементов песни, которые являются целью для выбора спаривания у этого вида (Bitterbaum & Baptista 1979; Mennill et al., 2006). Во-вторых, фоновые шумы городской среды больше всего влияют на вокализацию на максимальных частотах (Patricelli & Blickley, 2006; Slabbekoorn & Smith, 2002), которые, в свою очередь, больше всего подвержены изменениям клюва, характерным для городской среды. Наконец, мексиканские чечевицы имеют необычайно маленькие районы для пения - в некоторых местах менее 5 км (Bitterbaum & Baptista, 1 979; Tracy & Baker, 1999).
В целом, по всему спектру у городских чечевиц было меньше нот в их песнях и меньше трелей, чем у сельских (Badyaev et al., 2008). Взаимосвязь между морфологией клюва и характеристиками песни варьировалась в разных популяциях из-за различий в морфологии клюва (Badyaev & Hill, 2000), а также из-за местного культурного наследования песен. Например, в популяциях предков на юго-востоке Аризоны городские мексиканские чечевицы пели меньше нот и имели более медленные частоты трелей в более широком частотном диапазоне, чем вьюрки пустыни - различия, в основном, связаны с тем, что их клювы были длиннее, глубже и шире (Бадяев и др., 2008). Точно так же, в городской популяции на северо-западе Соединенных Штатов, чечевицы пели песни без жужжания - элемент песни, на который больше всего влияет городской шум (Mennill et al., 2006). Связь между экологическими условиями и сексуальной передачей сигналов посредством репродуктивной изоляции усиливает отбор для локальной адаптации в конфигурации клюва и, в конечном счете, способствует более быстрой дивергенции популяции.