Законы Мендаля
Законы Менделя — это принципы передачи наследственных признаков от родительских организмов к их потомкам,вытекающие из экспериментов Грегора Менделя. Эти принципы послужили основой для классической генетики и впоследствии были объяснены как следствие молекулярных механизмов наследственности. Хотя в русскоязычных учебниках обычно описывают три закона, «первый закон» не был открыт Менделем. Особое значение из открытых Менделем закономерностей имеет «гипотеза чистоты гамет»
I закон Менделя — закон единообразия первого поколения при скрещивании чистых форм.
Для примера можно рассмотреть этот процесс на кошках.
Произведем скрещивание двух гомозиготных по генам окраса кошек — черной и шоколадной.
В — аллель, отвечающий за синтез черного пигмента;
b — аллель, отвечающий за синтез шоколадного пигмента.
Черный кот имеет генотип ВВ, шоколадная кошка — bb. Родителей в генетике обозначают латинской буквой Р (от латинского parenta — «родители»). Сперматозоиды будут нести один аллель В, а яйцеклетки — аллель b.
В результате оплодотворения образуются зиготы, содержащие диплоидный набор хромосом (характерный набор для оплодотворенной яйцеклетки и всех соматических клеток гена) и несущие аллели Bb. Гибриды первого поколения, которые в генетике принято обозначать F1, окажутся гетерозиготными по данному локусу — Bb. Аллель В полностью доминирует над аллелем b, поэтому все полученные котята будут черными. Иногда доминирование одного аллеля над другим обозначают таким образом: В>b.
При скрещивании гомозиготных кошек получается одинаковое по фенотипу потомство. Эти результаты иллюстрируют I закон Менделя.
Анализируя данное скрещивание, мы говорим только об одном признаке — черном и шоколадном окрасе. Все многообразие признаков, определяющих как сходство, так и различие родителей, в данный момент нас не интересует. Такой тип скрещивания называют моногибридным.
II закон Менделя — закон расщепления: при скрещивании гибридов первого поколения между собой в потомстве второго поколения наблюдается закономерное расщепление.
Произведем скрещивание между собой потомков из первого поколения (F,).
Черные гетерозиготные кот и кошка, имеющие генотип Bb, образуют половые клетки двух типов, несущие аллель В и аллель b. При оплодотворении образуются следующие варианты зигот: BB Bb Bb bb в соотношении 1:2:1.
Во втором поколении (F2) получилось 3/4 черных котят и 1/4iшоколадных.
Черные гомозиготные и черные гетерозиготные котята выглядят одинаково — имеют одинаковый фенотип. В таком случае расщепление по фенотипу составит 3:1. Однако такое доминирование наблюдается не всегда. Известны и другие варианты.
III закон Менделя — аллели каждого гена распределяются в потомстве независимо от аллелей другого гена.
Для объяснения третьего закона Менделя используем для сходства скрещивания животных того же окраса, однако предположим, что у черного кота шерсть короткая, а у кошки — длинная. Длина шерсти определяется геном локуса Long, его доминанный аллель L определяет короткую, а рецессивный l — длинную шерсть. Опять-таки примем, что родители гомозиготны по исходным генам: генотип кота можно записать как BBLL, генотип кошки — bbll. Гены, определяющие эти признаки, относятся к разным локусам, а это значит, что у каждого из родителей будут образовываться половые клетки только одного типа.
Все котята первого поколения пойдут в отца — такие же черные и короткошерстные. И гетерозиготные по обоим генам — BbLl. А вот котята второго поколения (от скрещивания этих потомков исходной пары) будут уже четырех типов: больше всего — короткошерстных черных, примерно поровну — короткошерстных шоколадных и длинношерстных черных и совсем мало — длинношерстных шоколадных. Если посчитать значительное количество таких потомков (более 64), то получится примерное соотношение: 9 черных короткошерстных, 3 черных длинношерстных, 3 шоколадных короткошерстных, 1 шоколадный длинношерстный. То есть для каждого признака: окраса и длины шерсти соблюдается то же соотношение — 3:1 (12 черных, 4 шоколадных, 12 короткошерстных, 4 длинношёрстных). Получается, что признаки наследуются как бы независимо друг от друга.
Это всего лишь схема принципов «работы» генетики кошек и того, как создаются новые породы. На самом деле все не так уж просто.
Домина́нтность (доминирование) — форма взаимоотношений между аллелями одного гена, при которой один из них (доминантный) подавляет (маскирует) проявление другого (рецессивного) и таким образом определяет проявление признака как у доминантных гомозигот, так и у гетерозигот.
Рецесси́вный ген (англ. recessive gene) — генетическая информация, которая может подавляться воздействием доминантного гена и не проявляется в фенотипе. Рецессивный ген способен обеспечить проявление определяемого им признака только в том случае, если находится в паре с соответственным рецессивным геном. Если же он находится в паре с доминантным геном, то он не проявляется, так как доминантный ген подавляет его. Свойства, представленные рецессивными генами, проявляются в фенотипе у потомка лишь в том случае, если у обоих родителей присутствует рецессивный ген.
Полное доминирование. При полном доминировании фенотип гетерозиготы не отличается от фенотипа доминантной гомозиготы. Видимо, в чистом виде полное доминирование встречается крайне редко или не встречается вовсе. Например, люди, гетерозиготные по гену гемофилии А (сцепленный с Х-хромосомой рецессивный ген), имеют половинное количество нормального фактора свертывания по сравнению с гомозиготными по нормальному аллелю людьми, и активность фактора свертывания VIII у них в среднем вдвое ниже, чем у здоровых людей. В то же время у здоровых людей активность этого фактора варьирует от 40 до 300 % по сравнению со средней для популяции. Поэтому наблюдается значительное перекрывание признаков у здоровых и носителей-гетерозигот. При фенилкетонурии (аутосомно-рецессивный признак) гетерозиготы обычно считаются здоровыми, однако активность печёночного фермента фенилаланин-4-гидроксилазы у них вдвое ниже нормы, а содержание фенилаланина в клетках повышено, что, по некоторым данным, приводит к снижению IQ и повышенному риску развития некоторых психотических расстройств.
Неполное доминирование. При неполном доминировании гетерозиготы имеют фенотип, промежуточный между фенотипами доминантной и рецессивной гомозиготы. Например, при скрещивании чистых линий львиного зева и многих других видов цветковых растений с пурпурными и белыми цветками особи первого поколения имеют розовые цветки. При скрещивании чистых линий андалузских кур чёрной и белой окраски в первом поколении рождаются куры серой окраски. На молекулярном уровне самым простым объяснением неполного доминирования может быть как раз двукратное снижение активности фермента или другого белка (если доминантный аллель дает функциональный белок, а рецессивный — дефектный). Например, за белую окраску может отвечать дефектный аллель, который дает неактивный фермент, а за красную — нормальный аллель, который дает фермент, производящий красный пигмент. При половинной активности этого фермента у гетерозигот количество красного пигмента снижается вдвое, и окраска розовая. Могут существовать и другие механизмы неполного доминирования.
При неполном доминировании во втором поколении моногибридного скрещивания наблюдается одинаковое расщепление по генотипу и фенотипу в соотношении 1:2:1.
Кодоминирование. При кодоминировании, в отличие от неполного доминирования, у гетерозигот признаки, за которые отвечает каждый из аллелей, проявляются одновременно (смешанно). Типичный пример кодоминирования — наследование групп крови системы АВ0 у человека. Всё потомство людей с генотипами АА (вторая группа) и ВВ (третья группа) будет иметь генотип АВ (четвертая группа). Их фенотип не является промежуточным между фенотипами родителей, так как на поверхности эритроцитов присутствуют оба агглютиногена (А и В). При кодоминировании назвать один из аллелей доминантным, а другой — рецессивным нельзя, эти понятия теряют смысл: оба аллеля в равной степени влияют на фенотип. На уровне РНК и белковых продуктов генов, видимо, подавляющее большинство случаев аллельных взаимодействий генов — это кодоминирование, ведь каждый из двух аллелей у гетерозигот обычно кодирует РНК и/или белковый продукт, и оба белка или РНК присутствуют в организме.
Аллелизм — явление парности (или множественности) признаков организма, которые исключают друг друга. Возникает под влиянием аллельных генов.
Генотип и фенотип. Фенотип – это совокупность всех или анализируемых признаков, характерных для данного организма.
Фенотип развивается на основе внутренних факторов, материнских факторов и за счет влияния факторов внешней среды. К внутренним факторам относится наследственная информация, которую на уровне вида можно определить как геном, а на уровне организма понятием генотип (совокупность аллелей). Гены в организме представлены в определенном физикохимическом состоянии, которое определяется последовательностью нуклеотидов. Конкретной последовательностью нуклеотидов является аллель. Как правило для данного вида организмов характерны различные варианты аллелей, то есть фенотип определяется явление множественного аллелизма. Множественный аллелизм заключается в том, что различные аллели контролируют развитие более, чем двух вариантов конкретного признака (конкретный вариант признака называется феном). Причиной множественного аллелизма является внутригенная мутация, внутригенный кроссинговер. Классическим примером множественного аллелизма является наследование групп крови у человека. Гены в системе АВО отвечают за синтез различных галактозилтрансфераз. Множественный аллелизм характерен для окраски глаз у дрозофилы.
С abr – абрикосовый
C cr - коралловый
C + - красный
C ch - вишневый
C eor - эозинофиловый
C – белые
Множественный аллелизм характерен для окраски шерсти у многих млекопитающих. Существует ген С, которые контролирует структуру фермента тирозолазы, субстратом для этого фермента является тирозин, который под действием фермента превращается в пигмент меланин. Существует два варианта меланина:
Эумеланин
Феомеланин
Существуют различные аллели данного гена:
С - черный
C ch - шиншилловая
C h – гималайская
С – белый
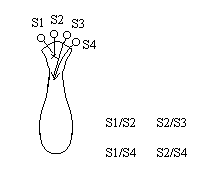
Множественный аллелизм лежит в основе генетического полиморфизма популяции. Множественный аллелизм существует у растений в основном у перекрестно опыляемых, причем существуют механизмы, которые поддерживают этот генетический полиморфизм. К таким механихмам относится явление самостерильности у покрытосемянных растений, у них существует ген S, который определяет проростание пыльцевой трубки.
Существует несколько аллелей генов S (S 1, S 2, S 3…).
Влияние генотипа на фенотип как правило очень сложное и один ген может влиять не на один, а на целую группу признаков. Такое явление получило название плейотропия или множественное действие генов. Различают два варианта плейотропии:
- первичная или структурная
- вторичная или функциональная
при первичной плейотропии патологичен продукт гена, но это влияет на множество признаков в организме. Синдром Марфана, болезнь, связанная с патологией в синтезе каллогена. В результате патологично большинство каллогеновых структур, в организме наблюдается деформация скелета, рост, как правило, выше среднего, непропорционально удлинены конечности, арахнодактелия, патологии суставных сумок, различные пороки сердца, патологии органов зрения.
Вторичная плейотропия. Классическим примеро является серповидноклеточная анемия. Эта аномалия вызвана одной аминокислотной заменой в В-цепи гемоглобина, в результате гемоглобин плохо связывается с кислородом и ткани накодятся в постоянном дефиците кислорода. Это доминантное заболевание, гомозиготы не выживают, у гетерозигогт наблюдается увеличение селезенки, гипертрофия сердца, гипертрофия красного костного мозга. Гетерозиготы оказываются в некоторых условиях более жизнеспособны, чем гомозиготы по рецессиву (устойчивость к малярии).
Патологичные гены не всегда проявляются в фенотипе, причем они могут проявляться с разной степенью выраженности. Вероятность проявления данного аллеля в фенотипе называется пенетрантностью. Если аллель всегда проявляется в фенотипе, то говорят о полной пенетрантности, если аллель не проявляется в фенотипе в различных случаях, то говорят о неполной пенетрантности. Например, полидактилия – это болезнь с неполной пенетрантностью. Из 100 носителей аллелей она проявляется в фенотипе с разной степенью выраженности. Например, полидактилия – это появление пятого пальца на всех конечностях, либо только на одной, в виде нормально развитого пальца, либо только в виде зачатка фаланги, в этом случае говорят о вариабельной экспрессивности развития признаков. Экспрессивность развития признаков бывает константной, если данный аллель всегда проявляется в фенотипе. Влияние генотипа на фенотип проявляется в явлении взаимодействия генов.
4. Наследственность и среда. Концепция нормы реакции и развитие. Морфогенез нервной системы и факторы, влияющие на этот процесс. Роль эмбрионального и неонального опыта в развитии поведения. Родительские эффекты в развитии. Эпигенез. Случайности развития. Историзм развития.
Наследственность и среда
Развитие поведения может быть предметом изучения педагогической, детской, дифференциальной, сравнительной психологии, психогенетики, а также биохимии, эмбриологии, генетики, физиологии, этологии.
Психобиология развития - молодая область науки. В которой происходит интеграция многих подходов.
Эпигенез. Развитие особи от зародыша до взрослого организма включает множество взаимодействий между генетическим аппаратом животного и его окружающей средой. Это взаимодействие предполагает, что каждая стадия развития служит основой для последующей. Этот процесс называется эпигенезом, который, в современном понимании, включает последовательное развертывание генетической информации при участии нейрогуморальных, гормональных и средовых факторов. Эпигенез представляет развитие как системный динамический процесс, в котором гены могут играть роль своеобразных триггеров в процессах дифференцировки клеток, напрвляющих развитие по тому или иному пути. При этом негенетические факторы (сигналы из окружающей среды и других клеточных систем) могут модифицировать развитие.
Для многих видов характерны периоды в развитии, когда особь чувствительна к определенным типам влияния извне. То, чему животное научается во время этого чувствительного периода, обычно сохраняется на протяжении всей его дальнейшей жизни.
Молодым животным часто свойственно особое, ювенильное поведение, которое утрачивается во взрослом состоянии.
Для поведения, которое имеет место без очевидного влияния окружающей среды, используется термин врожденное, однако, надо не забывать, что факторы окружающей среды в какой-то степени влияют на все поведение в целом.
В некоторых случаях можно выявить действие одного гена на поведение, но обычно даже простые аспекты поведения контролируются множеством генов (полигенное наследование).
В трудах отечественных исследователей показано, что в онтогенезе происходят не только количественные, но и качественные изменения психических функций: меняется их структура, взаимосвязи и иерархия, и, главное, меняются механизмы их реализации (Выготский, 1960; Лурия, 1962). В развитии происходит "смена его детерминант, а вместе с тем и смена системных оснований психических качеств (Ломов, 1984).
В плане психогенетическом это означает, что, оставаясь фенотипически тем же, психологический признак, меняя в процессе развития реальное психологическое содержание, может менять и отношение к генотипу (Лурия, 1962).
Это не означает, что с возрастом происходит, например, автоматическое накопление средовых влияний, как и не означает простой замены элементарных форм функций высшими, социально опосредованными. У взрослого человека эти формы сосуществуют.
Все сказанное позволяет сформулировать совсем иной взгляд на роль генотипа и среды в формировании функции: генетически "плохой" признак (например, плохая образная память) в онтогенезе или через специальные воздействия может быть переведен на другой уровень реализации и тем самым "улучшен". Возможно и другое: в разных возрастах само выполнение психодиагностических заданий может происходить по разным основаниям, в него включаются разные психологические качества.
Безусловно, те психологические признаки, в изменчивости которых генетические детерминанты надежно установлены, относятся к капитальным, конституциональным особенностям человека. Однако необходимо, в частности, создать типологию индивидуального развития, выявить роль генотипических и средовых факторов в его детерминации, прежде чем это знание можно будет с уверенностью рекомендовать практике.
Генетика поведения человека лишена возможности экспериментировать с объектом своего исследования, поэтому изучение развития ведется также на популяционном уровне. Основная часть выводов генетики поведения носит статистический характер, поскольку касается не реального взаимодействия генотипа и среды в процессе развития конкретного фенотипа, а имеет отношение к компонентам дисперсии фенотипов в популяции. Когда на основании коррелирующего поведения у родственников популяционные генетики делают вывод о наследуемости признака, это не означает, что индивидуальное развитие данного поведения обусловлено исключительно генетическими причинами. Высокая наследуемость свидетельствует лишь о том, что разнообразие индивидов в популяции в значительной степени связано с генотипическими различиями между ними.