Иммунная система - это специализированная, анатомически обособленная лимфоидная ткань. Она распределена по всему организму в виде различных лимфоидных образований и отдельных клеток, и на ее долю приходится 1-2% от массы тела. В анатомическом плане иммунная система подразделена на центральные и периферические органы, в функциональном - на органы воспроизводства и селекции клеток (костный мозг, тимус), контроля внешней среды или экзогенной интервенции (лимфоидные системы кожи и слизистых оболочек), контроля генетического постоянства внутренней среды (селезенка, лимфатические узлы, печень, кровь, лимфа).
Основными функциональными клетками являются лимфоциты. Их количество в организме достигает 1012. К числу функциональных клеток иммунной системы относят также мононуклеарные и гранулярные лейкоциты, тучные и дендритные клетки. Часть клеток сосредоточена в отдельных органах иммунной системы, другие свободно перемещаются по всему организму. Схематическое строение иммунной системы представлено на рис. 10.3.
10.2.1.1. Центральные органы иммунной системы
Центральные органы иммунной системы, костный мозг и вилочковая железа или тимус, - это органы воспроизводства и селекции клеток иммунной системы. Здесь происходят лимфопоэз - рождение, размножение (пролиферация) и дифференцировка лимфоцитов до стадии предшественников или зрелых неиммунных (наивных) клеток, а также их «обучение». У птиц к центральным органам иммунной системы относят сумку Фабрициуса (bursaFabricii), локализованную в области клоаки.
Костный мозг располагается в губчатом веществе костей (эпифизы трубчатых костей, грудина, ребра и др.). Здесь находятся полипотентные стволовые клетки (ППСК), которые являются ро-
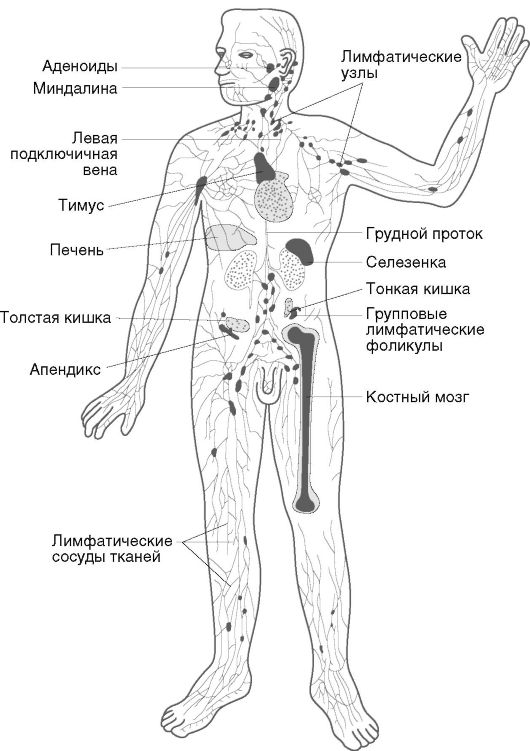 Рис. 10.3. Органы иммунной системы человека
Рис. 10.3. Органы иммунной системы человека
доначальницами всех форменных элементов крови, включая иммунокомпетентные клетки. В строме костного мозга формируются предшественники В- и Т-лимфоцитов, которые впоследствии мигрируют соответственно в В-зоны макроорганизма и тимус. Фагоциты и некоторые дендритные клетки также образуются в костном мозгу. В нем можно также обнаружить плазматические клетки - результат терминальной дифференцировки В-лимфоцитов.
Вилочковая железа, тимус, или зобная железа, располагается в верхней части загрудинного пространства. Этот орган отличается особым морфогенезом. Тимус формируется в период внутриутробного развития. К моменту рождения масса тимуса достигает 10-15 г, окончательно он созревает к пятилетнему возрасту, а максимального размера достигает к 10-12 годам жизни (масса 30-40 г). После периода полового созревания начинается инволюция органа - происходит замещение лимфоидной ткани жировой и соединительной.
Тимус имеет дольчатое строение. В его структуре различают мозговой и корковый слои. В строме коркового слоя находится большое количество эпителиальных клеток коры, названных «клеткиняньки», которые своими отростками образуют мелкоячеистую сеть, где располагаются созревающие лимфоциты. В пограничном, корково-мозговом, слое располагаются дендритные клетки тимуса, а в мозговом - эпителиальные клетки мозгового слоя.
Предшественники Т-лимфоцитов поступают из костного мозга в корковый слой тимуса. Здесь под влиянием тимических факторов они активно размножаются, дифференцируются (превращаются) в зрелые Т-лимфоциты и «учатся» распознавать чужеродные антигенные детерминанты.
Процесс «обучения» включает положительную и отрицательную селекцию. Критерием «обученности» являются качество Т-клеточной антигенной рецепции (специфичность и аффинность) и жизнеспособность клетки.
Положительная селекция происходит в корковом слое при помощи эпителиальных клеток. Суть ее заключается в поддержке клонов Т-лимфоцитов, рецепторы которых эффективно связались с экспрессированными на эпителиальных клетках молекулами MHC, независимо от структуры инкорпорированных собственных олигопептидов. Эпителиоциты коры выделяют ростовые факторы тимуса, активирующие размножение Т-лимфоцитов.
Отрицательную селекцию осуществляют дендритные клетки в пограничной корково-мозговой зоне тимуса. Ее цель - выбраковка аутореактивных клонов Т-лимфоцитов. Клетки, позитивно реагирующие на комплекс MHC-аутологичный пептид, подвергаются уничтожению путем индукции у них апоптоза.
В итоге селекции более 99% Т-лимфоцитов не выдерживают испытаний и погибают. Лишь менее 1% клеток превращается в зрелые формы, способные распознать в комплексе с аутологичными MHC только чужеродные биополимеры. Ежесуточно около 106 зрелых «обученных» Т-лимфоцитов покидают тимус с крово- и лимфотоком и мигрируют в различные органы и ткани.
Созревание и «обучение» Т-лимфоцитов в тимусе имеет важное значение для формирования иммунитета. Отсутствие или недоразвитие тимуса при врожденном дефекте развития вилочковой железы - аплазии или гипоплазии органа, ее хирургическом удалении или радиационном поражении ведет к резкому снижению эффективности иммунной защиты макроорганизма. Между тем тимэктомия у взрослых практически не приводит к серьезным дефектам в иммунитете.
10.2.1.2. Периферические органы иммунной системы
К периферическим органам иммунной системы относят селезенку, лимфатические узлы, аппендикс, печень, миндалины глоточного кольца, групповые лимфатические фолликулы, кровь, лимфу и др. В этих органах проходит иммуногенез - размножение и окончательное созревание предшественников иммунокомпетентных клеток и осуществляется иммунологический надзор. В функциональном плане периферические органы иммунной системы могут быть подразделены на органы контроля внутренней среды организма (лимфатические узлы, селезенка, тканевые мигрирующие клетки) и его кожных и слизистых покровов (аппендикс, лимфатические фолликулы и скопления).
Лимфатические узлы - мелкие округлые анатомические образования бобовидной формы, которые располагаются по ходу лимфатических сосудов. Каждый участок тела имеет региональные лимфоузлы. В общей сложности в организме человека насчитывается до 1000 лимфоузлов. Лимфатические узлы выполняют функцию биологического сита - через них фильтруется лимфа и задерживаются и концентрируются антигены. Через лимфоузел проходит в среднем около 109 лимфоцитов в 1 ч.
В строении лимфоузла различают корковое и мозговое вещество. Строма коры разделена соединительнотканными трабекулами на сектора. В ней выделяют поверхностный корковый слой и паракортикальную зону. В секторах поверхностного коркового слоя расположены лимфатические фолликулы с центрами размножения В-лимфоцитов (герминативные центры). Здесь же обнаруживаются фолликулярные дендритные клетки, способствующие созреванию В-лимфоцитов. Паракортикальный слой - это зона Т-лимфоцитов и интердигитальных дендритных клеток, потомков дермальных клеток Лангерганса. Мозговое вещество образовано тяжами соединительной ткани, между которыми располагаются макрофаги и плазматические клетки.
В пределах лимфоузла происходит антигенная стимуляция иммунокомпетентных клеток и включается система специфического иммунного реагирования, направленная на обезвреживание антигена.
Селезенка - это орган, через который фильтруется вся кровь. Он располагается в левой подвздошной области и имеет дольчатое строение. Лимфоидная ткань образует белую пульпу. В строении различают первичные, периартериальные лимфоидные фолликулы (окружают артерии по их ходу) и вторичные, располагающиеся на границах первичных фолликулов. Первичные лимфоидные скопления заселены преимущественно Т-лимфоцитами, а вторичные - В-лимфоцитами и плазматическими клетками. Кроме того, в строме селезенки обнаруживают фагоциты и ретикулярные дендритные клетки.
В селезенке, как в сите, задерживаются антигены, оказавшиеся в кровотоке, и состарившиеся эритроциты. Этот орган называют кладбищем эритроцитов. Здесь происходят антигенная стимуляция иммунокомпетентных клеток, развитие специфической иммунной реакции на антиген и его обезвреживание.
Печень играет особую роль в иммунной системе. В ней находится более половины всех тканевых макрофагов и большая часть естественных киллеров. Лимфоидные популяции печени обеспечивают толерантность к пищевым антигенам, а макрофаги утилизируют иммунные комплексы, в том числе сорбированные на стареющих эритроцитах.
Групповые лимфатические фолликулы (пейеровы бляшки) являются скоплением лимфоидной ткани в слизистой оболочке тонкой кишки. Такие образования также находятся в червеобразном отростке слепой кишки - аппендиксе. Кроме того, на всем протяже-
нии желудочно-кишечного тракта, начиная с пищевода и кончая анальным отверстием, располагаются единичные лимфатические фолликулы. Они обеспечивают местный иммунитет слизистой оболочки кишки и ее просвета и регулируют видовой и количественный состав ее нормальной микрофлоры.
Скопление лимфоидных элементов в виде миндалин глоточного кольца обеспечивает местный иммунитет в носоглотке, ротовой полости и верхних дыхательных путях, защищает их слизистые оболочки от внедрения микробов и других генетически чужеродных агентов, передающихся воздушно-капельным или воздушнопылевым путем, и регулирует локальную нормофлору.
Лимфа - жидкая ткань организма, которая содержится в лимфатических сосудах и узлах. Она включает в себя все соединения, поступающие из межтканевой жидкости. Основными и практически единственными клетками лимфы являются лимфоциты. В ее составе эти клетки осуществляют кругооборот в организме.
В кровь циркулируют предшественники и зрелые Т- и В-лимфоциты, полиморфно-ядерные лейкоциты, моноциты. Лимфоциты составляют 30% общего количества лейкоцитов. Одномоментно в крови присутствует менее 2% общего количества лимфоцитов.
10.2.1.3. Клетки иммунной системы
Специфическую функцию иммунной защиты непосредственно осуществляет многочисленный пул клеток миелоидного и лимфоидного ростков крови: лимфоциты, фагоциты и дендритные клетки. Это основные клетки иммунной системы. Кроме них, в иммунный ответ может вовлекаться множество других клеточных популяций (эпителий, эндотелий, фибробласты и др.). Перечисленные клетки различаются морфологически, по функциональной активности, маркерам (специфические молекулярные метки), рецепторному аппарату и продуктам биосинтеза. Тем не менее большую часть клеток иммунной системы объединяет близкое генетическое родство: они имеют общего предшественника, полипотентную стволовую клетку костного мозга (рис. 10.4).
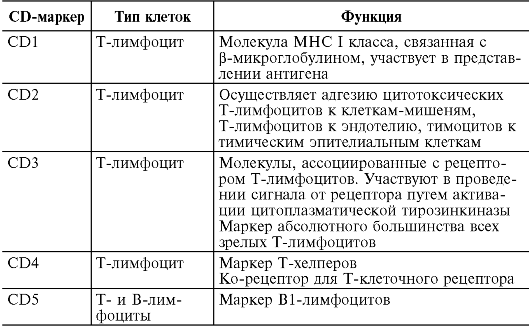
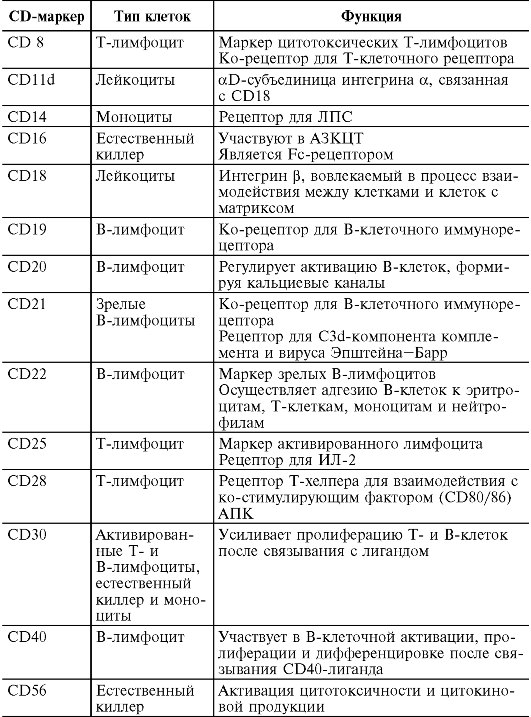
На поверхности цитоплазматической мембраны клеток иммунной системы существуют особые молекулы, которые служат их маркерами. В 80-х годах прошлого века была принята международная номенклатура мембранных маркеров лейкоцитов человека, названных «CD-антигены» (табл. 10.2)
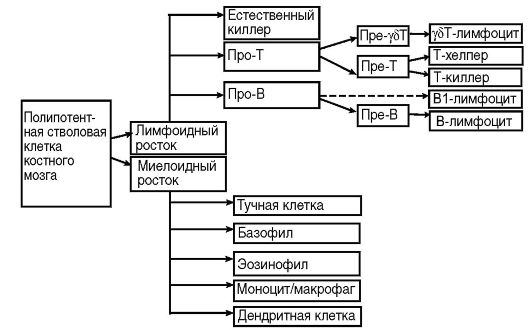 Рис. 10.4. Схема иммуногенеза (пояснения в тексте)
Рис. 10.4. Схема иммуногенеза (пояснения в тексте)
Таблица 10.2. Основные CD-маркеры клеток, участвующих в иммунном ответе
 Продолжение табл. 10.2
Продолжение табл. 10.2
 Окончание табл. 10.2
Окончание табл. 10.2
 Примечание. АЗКЦТ - антителозависимая клеточно-опосредованная цитотоксичность; АПК - антигенпрезентирующие клетки.
Примечание. АЗКЦТ - антителозависимая клеточно-опосредованная цитотоксичность; АПК - антигенпрезентирующие клетки.
По функциональной активности клетки-участники иммунного ответа подразделяют на регуляторные (индукторные), эффекторные и антигенпрезентирующие. Регуляторные клетки управляют функционированием компонентов иммунной системы путем выработки медиаторов - иммуноцитокинов и лигандов. Эти клетки определяют направление развития иммунного реагирования, его интенсивность и продолжительность. Эффекторы являются непосредственными исполнителями иммунной защиты путем прямого воздействия на объект либо путем биосинтеза биологически активных веществ со специфическим эффектом (антитела, токсичные субстанции, медиаторы и пр.).
Антигенпрезентирующие клетки выполняют ответственную задачу: захватывают, процессируют (перерабатывают путем ограниченного протеолиза) и представляют антиген иммунокомпетентным Т-клеткам в составе комплекса с MHC II класса. АПК лишены специфичности в отношении самого антигена. Молекула MHC II класса может включать в себя любые эндоцитированные из межклеточной среды олигопептиды, как свои собственные, так и чужие. Установлено, что большая часть комплексов MHC II класса содержит аутогенные молекулы и лишь малая доля - чужеродный материал.
Помимо MHC II класса, АПК экспрессируют ко-стимулирующие факторы (CD40, 80, 86) и множество молекул адгезии. Последние обеспечивают тесный, пространственно стабильный и продолжительный контакт АПК с Т-хелпером. Кроме того, АПК экспрессируют молекулы CD1, с помощью которых могут презентироватьлипосодержащие или полисахаридные антигены.
Основными профессиональными АПК являются дендритные клетки костно-мозгового происхождения, В-лимфоциты и макро-
фаги. Дендритные клетки почти в 100 раз эффективнее макрофагов. Функцию непрофессиональных АПК могут также выполнять некоторые другие клетки в состоянии активации - эпителиальные клетки и эндотелиоциты.
Осуществление целенаправленной иммунной защиты макроорганизма возможно благодаря наличию на клетках иммунной системы специфических антигенных рецепторов (иммунорецепторов). По механизму функционирования они подразделяются на прямые и непрямые. Прямые иммунорецепторы непосредственно связываются с молекулой антигена. Непрямые иммунорецепторы взаимодействуют с молекулой антигена опосредованно - через Fc-фрагмент молекулы иммуноглобулина (см. раздел 11.1.2). Это так называемый Fc-рецептор (FcR).
Fc-рецепторы различаются по аффинности. Высокоаффинный рецептор может связываться с интактными молекулами IgE или IgG4 и образовывать рецепторный комплекс, в котором антигенспецифическую ко-рецепторную функцию выполняет молекула иммуноглобулина. Такой рецептор есть у базофилов и тучных клеток. Низкоаффинный FcR распознает молекулы иммуноглобулина, уже образовавшие иммунные комплексы. Он обнаруживается на макрофагах, естественных киллерах, эпителиальных, дендритных и множестве других клеток.
Иммунное реагирование основано на тесном взаимодействии различных клеточных популяций. Это достигается при помощи биосинтеза клетками иммунной системы широкого спектра иммуноцитокинов. Подавляющее большинство клеток иммунной системы постоянно перемещается во внутренних средах организма с крово- и лимфотоком и благодаря амебоидной подвижности.
Клеточно-элементный состав иммунной системы постоянно возобновляется за счет деления стволовых клеток. Состарившиеся, выработавшие свой биологический ресурс, ложно активированные, зараженные и генетически трансформированные клетки уничтожаются.
10.2.1.3.1. Лимфоциты
Лимфоциты - подвижные мононуклеарные клетки. В зависимости от места созревания эти клетки подразделяются на две популяции Т- (тимус) и В- (бурса Фабрициуса, костный мозг) лимфоцитов. Лимфоциты играют ключевую роль в обеспечении приобретенного (адаптивного) иммунитета. Они осуществляют
специфическое распознавание антигена, индукцию клеточного и гуморального иммунного ответа, различные формы иммунного реагирования.
В организме непрерывно идет возобновление популяций лимфоцитов, клетки активно мигрируют между различными органами и тканями. Вместе с тем миграция и расселение лимфоцитов в тканях не являются хаотическим процессом. Он имеет направленный характер и строго регулируется экспрессией на мембране лимфоцитов, эндотелии сосудов и клеточных элементах стромы особых молекул адгезии (интегрины, селектины и др.). Так, незрелые Т-лимфоциты активно мигрируют в тимус. Зрелые неиммунные («наивные») лимфоциты тропны к периферическим лимфоидным органам и тканям. При этом Т- и В-лимфоциты заселяют только «свои» области - это так называемый эффект хоминговой рецепции (от англ. home - дом). Зрелые иммунные (активированные) лимфоциты распознают эпителий в очаге воспаления. Клетки иммунологической памяти всегда возвращаются в места своего происхождения.
Продолжительность жизни неиммунных лимфоцитов достаточно большая. У Т-лимфоцитов она достигает нескольких месяцев или лет, а у В-клеток - недель или месяцев. Дольше всех живут клетки иммунологической памяти (см. раздел 11.5) - до 10 лет и более. Однако активированные или терминально дифференцированные лимфоциты имеют короткую продолжительность жизни (несколько суток). Состарившиеся, ложно активированные и аутореактивные (реагирующие на аутоантигены) лимфоциты подвергаются уничтожению путем индукции у них апоптоза. Погибшие лимфоциты постоянно заменяются новыми за счет их пролиферации в центральных и периферических органах иммунной системы. Численность лимфоидных популяций находится под жестким контролем клеток самой иммунной системы.
Для выполнения специфической функции лимфоциты несут на своей поверхности прямые антигенные рецепторы и являются иммунокомпетентными клетками. Иммунорецептор В-лимфоцита и особого γδТ-лимфоцита распознает нативныйэпитоп, т.е. непосредственно отличает чужеродные субстанции. Иммунорецептор традиционного Т-лимфоцита ориентирован на олигопептиды в составе MHC, т.е. распознает измененное «свое».
Антигенспецифические рецепторы лимфоцитов имеют сложное молекулярное строение, уникальное для каждой клетки. Напри-
мер, у Т-лимфоцитов они состоят из нескольких полипептидных субъединиц, имеющих полигенное кодирование. Число генов, детерминирующих структуру V-области этого рецептора (вариабельный участок, ответственный за специфическое распознавание), в незрелой клетке достигает 100. При созревании лимфоцита в результате рекомбинационных перестроек в V-генах, индивидуальных для каждой клетки, образуется бесконечно большое количество вариантов антигенной специфичности рецептора, достигающее 1012, что сопоставимо с общей численностью популяции Т-лимфоцитов. Формирование В-клеточного рецептора имеет те же закономерности. Биологический смысл феномена чрезвычайно важен: в организме постоянно поддерживается широкий репертуар специфической направленности лимфоидных рецепторов, и клетки готовы в любой момент ответить защитной реакцией на любой возможный антиген.
В такой ситуации закономерно появление Т-лимфоцитов, специфичных для антигенов собственного организма. Однако они должны элиминироваться в тимусе на ранних этапах своего развития. Поэтому различают первичный и вторичный антигенраспознающий репертуар лимфоидных популяций. Первичный характеризуется набором рецепторных специфичностей, формирующимся при образовании лимфоцитов в костном мозгу индивидуума. Вторичный, или клональный, репертуар является совокупностью вариантов рецептора после отбраковки аутореактивных клонов клеток.
Антигенспецифическая рецепция в лимфоцитах имеет стандартные механизмы реализации. Полученный внеклеточной частью рецептора сигнал от раздражителя (антигена) передается по трансмембранному участку на его внутриклеточную часть, которая уже активирует внутриклеточные ферменты (тирозинкиназу, фосфорилазу и др.).
Для запуска продуктивной реакции лимфоцита необходима агрегация его рецепторов. Кроме того, для стабилизации рецепторлигандного взаимодействия и восприятия ко-стимулирующего сигнала требуются вспомогательные молекулы.
Среди лимфоцитов встречаются клетки без отличительных признаков Т- и В-лимфоцитов. Они получили название нулевых клеток. В костном мозгу на их долю приходится около 50% всех лимфоцитов, а в крови - примерно 5%. Функциональная активность остается неясной.
В-лимфоциты. В-лимфоциты - это преимущественно эффекторные иммунокомпетентные клетки, на долю которых приходится около 15% всей численности лимфоцитов. Выделяют две субпопуляции В-лимфоцитов: традиционные В-клетки, не имеющие маркера CD5-, и CD5+ В1-лимфоциты.
При электронной микроскопии CD5- В-лимфоциты имеют шероховатую поверхность, на ней определяются CD19-22 и некоторые другие. Функцию антигенспецифического рецептора (BCR) выполняют особые мембранные формы иммуноглобулинов. Клетки экспрессируют MHC II класса, ко-стимулирующие молекулы CD40, 80, 86, FcR к иммунным комплексам и нативным молекулам иммуноглобулина класса G, рецептор к эритроцитам мыши, иммуноцитокинам и др.
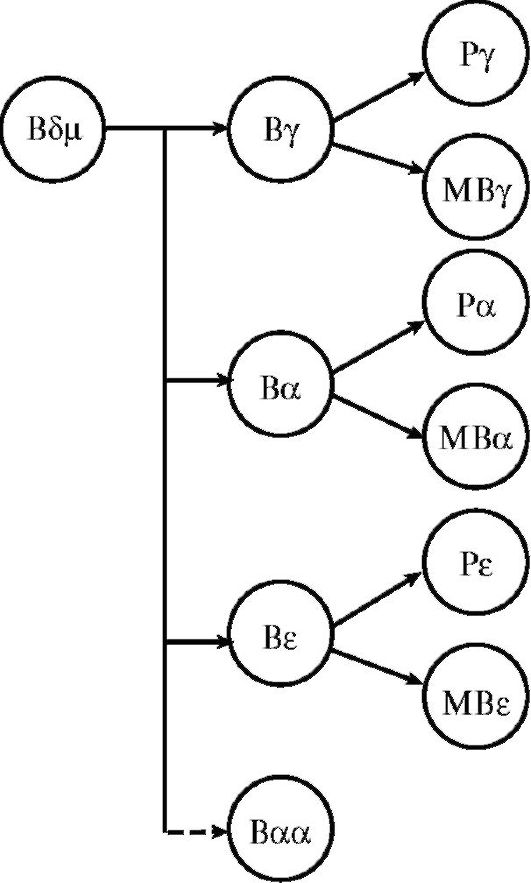 Рис. 10.5. Схема дифференцировки В-лимфоцита: Р - плазматическая клетка; МВ - В-лимфоцит иммунологической памяти; Вαα - синтезирует полимерный иммуноглобулин А в слизистых оболочках
Рис. 10.5. Схема дифференцировки В-лимфоцита: Р - плазматическая клетка; МВ - В-лимфоцит иммунологической памяти; Вαα - синтезирует полимерный иммуноглобулин А в слизистых оболочках
Функцией зрелых CD5- В-лимфоцитов и их потомков (плазмоцитов) является продукция иммуноглобулинов. Кроме того, В-лимфоциты являются профессиональными АПК. Они участвуют в формировании гуморального иммунитета, В-клеточной иммунологической памяти и гиперчувствительности немедленного типа.
Дифференцировка и созревание В-лимфоцитов (рис. 10.5) происходят сначала в костном мозге, а затем в периферических органах иммунной системы, куда они отселяются на стадии предшественников. Потомками В-лимфоцитов являются клетки иммунологической памяти и плазматические клетки. Основные морфологические признаки последних - развитый эндоплазматический ретикулум и аппарат Гольджи с большим количеством рибо-
сом. Плазмоцит имеет короткий период жизни - не более 2-3 сут.
В1-лимфоциты считают филогенетически наиболее древней ветвью антителопродуцирующих клеток. Предшественники этих клеток рано мигрируют в ткани слизистых оболочек, где автономно от центральных органов иммунной системы поддерживают численность своей популяции. Клетки экспрессируют CD5, синтезируют низкоаффинныеIgA и IgM к полисахаридным и липидным антигенам микробов и обеспечивают иммунную защиту слизистых оболочек от условно-патогенных бактерий.
Функциональной активностью В-лимфоцитов управляют молекулярные антигены и иммуноцитокины Т-хелпера, макрофага и других клеток.
Т-лимфоциты. Т-лимфоциты - это сложная по составу группа клеток, которая происходит от полипотентной стволовой клетки костного мозга, а созревает и дифференцируется в тимусе из предшественников. На долю этих клеток приходится около 75% всей лимфоидной популяции. На электронограмме все Т-лимфоциты имеют гладкую поверхность, их общим маркером являются CD3, а также рецептор к эритроцитам барана. В зависимости от строения антигенного рецептора (TCR) и функциональной направленности сообщество Т-лимфоцитов может быть разделено на группы.
Различают два типа TCR: αβ и γδ. Первый тип - гетеродимер, который состоит из двух полипептидных цепей - α и β. Он характерен для традиционных Т-лимфоцитов, известных как Т-хелперы и Т-киллеры. Второй обнаруживается на поверхности особой популяции γδТ-лимфоцитов.
Т-лимфоциты функционально также разделяются на две субпопуляции: иммунорегуляторов и эффекторов. Задачу регуляции иммунного ответа выполняют Т-хелперы. Ранее предполагалось существование Т-супрессоров, способных тормозить развитие иммунной реакции (супрессия). Однако до сих пор клетка морфологически не идентифицирована, хотя сам супрессорный эффект существует. Эффекторную функцию осуществляют цитотоксические лимфоциты Т-киллеры.
В организме Т-лимфоциты обеспечивают клеточные формы иммунного ответа (гиперчувствительность замедленного типа, трансплантационный иммунитет и т.д.), определяют силу и продолжительность иммунной реакции. Их созреванием, дифференцировкой и активностью управляют цитокины и макрофаги.
Т-хелперы. Т-хелперы или Т-помощники - субпопуляция Т-лимфоцитов, которые выполняют регуляторную функцию. На их долю приходится около 75% всей популяции Т-лимфоцитов. Они несут маркер CD4, а также αβ TCR, с помощью которого анализируют природу антигена, представляемую им АПК.
Рецепция антигена Т-хелпером, т.е. анализ его чужеродности, - весьма сложный процесс, требующий высокой точности. Ему способствуют (рис. 10.6) молекула CD3 (комплексируется с TCR), ко-рецепторные молекулы CD4 (имеют сродство к молекулярному комплексу MHC II класса), молекулы адгезии (стабилизируют межклеточный контакт), рецепторы (взаимодействуют с ко-стимулирующими факторами АПК - CD28, 40L).
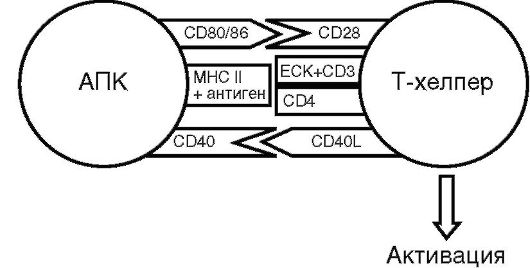 Рис. 10.6. Схема активации Т-хелпера (пояснение в тексте)
Рис. 10.6. Схема активации Т-хелпера (пояснение в тексте)
Активированный Т-хелпер продуцирует широкий спектр иммуноцитокитов, при помощи которых он управляет биологической активностью множества клеток, вовлеченных в иммунный ответ.
Популяция Т-хелперов гетерогенна. Активированный CD4+ Т-лимфоцит (ТΩ-хелпер) дифференцируется в одного из своих потомков: T1- или Т2-хелпер (рис. 10.7). Эта дифференцировка является альтернативной и направляемой цитокинами. Т1- или Т2-хелперы различаются лишь функционально по спектру продуцируемых цитокинов.
Т1-хелпер образует ИЛ-2, 3, γ-ИФН, ФНО и др., необходимые для развития клеточного иммунного ответа, гиперчувствительности замедленного типа, иммунного воспаления. Формирование этой клетки определяют активированный макрофаг, естественный и Т-киллеры, синтезирующие ИЛ-12 и γ-ИФН.
Т2-хелпер продуцирует ИЛ-4, 5, 6, 9, 10, 13 и др., которые поддерживают гуморальный иммунный ответ, а также гиперчув-
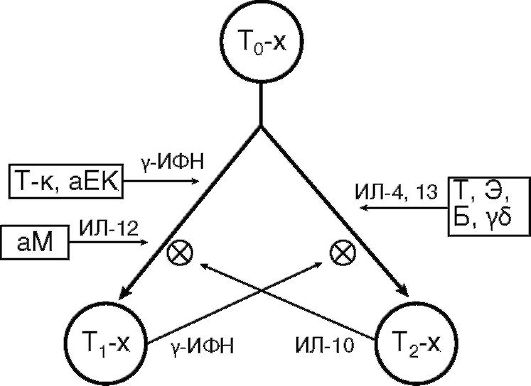 Рис. 10.7. Схема дифференцировки Т-хелпера: Т-х - Т-хелпер; аМ - активированный макрофаг; Т-к - Т-киллер; аЕК - активированный естественный киллер; Э - эозинофил; Б - базофил; Т - тучная клетка; γδТ - γδТ-лимфоцит
Рис. 10.7. Схема дифференцировки Т-хелпера: Т-х - Т-хелпер; аМ - активированный макрофаг; Т-к - Т-киллер; аЕК - активированный естественный киллер; Э - эозинофил; Б - базофил; Т - тучная клетка; γδТ - γδТ-лимфоцит
ствительность немедленного типа. Дифференцировку в сторону Т2-хелпера потенцируют γδТ-клетки, базофилы, тучные клетки и эозинофилы, синтезирующие ИЛ-4 и 13.
В организме поддерживается баланс Т1-/Т2-хелперов, который необходим для развития адекватного иммунного ответа. Т1- и Т2- хелперы являются антагонистами и тормозят развитие друг друга. Установлено, что в организме новорожденных преобладают Т2-хелперы. Нарушение заселения желудочно-кишечного тракта нормальной микрофлорой тормозит развитие субпопуляции Т1- хелперов и ведет к аллергизации организма.
Т-киллеры (цитотоксические Т-лимфоциты). Т-киллер - субпопуляция Т-лимфоцитов-эффекторов, на долю которых приходится примерно 25% всех Т-лимфоцитов. На поверхности Т-киллера определяются молекулы CD8, а также αβ TCR к антигену в комплексе с MHC I класса, по которому «свои» клетки отличаются от «чужих». В рецепции принимают участие молекула CD3, комплексирующая с TCR, и ко-рецепторные молекулы CD8, тропные к MHC I класса (рис. 10.8).
Т-киллер анализирует клетки собственного организма в поисках чужеродного MHC I класса. Клетки мутантные, пораженные вирусом, или аллогенного трансплантата несут на своей поверхности такие признаки генетической чужеродности, поэтому являются мишенью Т-киллера.
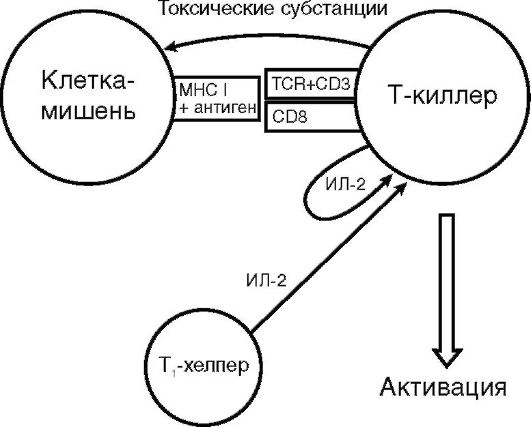 Рис. 10.8. Схема активации Т-киллера (пояснения в тексте)
Рис. 10.8. Схема активации Т-киллера (пояснения в тексте)
Т-киллер устраняет клетки-мишени путем антителонезависимой клеточно-опосредованной цитотоксичности (АНКЦТ) (см. раздел 11.3.2), для чего синтезирует ряд токсичных субстанций: перфорин, гранзимы и гранулизин. Перфорин - токсичный белок, который синтезируют цитотоксические лимфоциты-Т-киллеры и естественные киллеры. Обладает неспецифическим свойством. Вырабатывается только зрелыми активированными клетками. Перфорин образуется в виде растворимого белка-предшественника и накапливается в цитоплазме в гранулах, которые сосредоточиваются около TCR, связавшегося с клеткой-мишенью для обеспечения локального, адресного поражения клетки-мишени. Содержимое гранул высвобождается в узкую синаптическую щель, образованную тесным контактом цитотоксического лимфоцита и клеткимишени. За счет гидрофобных участков перфорин встраивается в цитоплазматическую мембрану клетки-мишени, где в присутствии ионов Са2+ полимеризуется в трансмембранную пору диаметром 16 нм. Образовавшийся канал может вызвать осмотический лизис клетки-мишени (некроз) и/или обеспечить проникновение в нее гранзимов и гранулизина.
Гранзимы - это обобщающее название сериновых протеаз, синтезируемых зрелыми активированными цитотоксическими лимфоцитами. Различают три типа гранзимов: А, В и С. После синтеза гранзимы накапливаются в гранулах подобно перфорину и вместе
с ним выделяются из клетки в синаптическую щель. В клеткумишень проникают через поры, образованные перфорином. Мишенью для гранзимов являются внутриклеточные ферменты, инициирующие апоптоз и обладающие широкой нуклеазной активностью, в том числе способностью разрушать нуклеиновые кислоты внутриклеточных паразитов. Таким образом, гранзимы индуцируют гибель клетки путем апоптоза и санацию организма от зараженных клеток.
Гранулизин - эффекторная молекула с ферментативной активностью, синтезируемая цитотоксическими лимфоцитами. Способен запускать в клетках-мишенях апоптоз, повреждая мембрану их митохондрий.
Т-киллер обладает огромным биологическим потенциалом - его называют серийным убийцем. За короткий срок он может уничтожить несколько клеток-мишеней, затрачивая на каждую около 5 мин. Эффекторную функцию Т-киллера стимулирует Т1-хелпер, хотя в ряде случаев его помощь не требуется. Помимо эффекторной функции, активированный Т-киллер синтезирует γ-ИФН и ФНО, стимулирующие макрофаг и потенцирующие иммунное воспаление.
γδ Т-лимфоциты. Среди Т-лимфоцитов существует малочисленная популяция клеток с фенотипом CD4-CD8-, которые несут на своей поверхности особый TCR γδ-типа - γδТ-лимфоциты. Локализуются в эпидермисе и слизистой оболочке желудочнокишечного тракта. Их общая численность не превышает 1% общего пула Т-лимфоцитов, однако в покровных тканях она может достигать 10%.
γδТ-лимфоциты происходят из автономного ростка стволовых клеток, мигрировавших в покровные ткани на ранних этапах эмбриогенеза. В созревании минуют тимус. Активируются клетками поврежденного эпителия желудочно-кишечного тракта и эпидермиса, размножение усиливается ИЛ-7.
Антигенный рецептор γδТ-лимфоцита сходен с BCR, его активный центр непосредственно связывается с эпитопом антигена без его предварительного процессинга и участия MHC. Антигенные детерминанты могут быть представлены, например, молекулами CD1. γδTCRориентированы на распознавание некоторых широко распространенных микробных антигенов (липопротеинов, белков теплового шока, бактериальных суперантигенов и др.).
γδТ-лимфоциты могут быть как эффекторными, цитотоксическими клетками (принимают участие в удалении патогенов на ранних этапах антиинфекционной защиты), так и регуляторами иммунореактивности. Синтезируют цитокины, активирующие местный иммунитет и локальную воспалительную реакцию, в том числе усиливают образование Т2-хелперов. Кроме того, γδ-клетки продуцируют ИЛ-7 и управляют численностью собственной популяции.
10.2.1.3.2. Другие клетки иммунной системы
Помимо лимфоцитов, в развитии иммунного ответа участвует множество различных клеточных популяций, относящихся в основном к миелоидному ростку. Особого внимания заслуживают естественные киллеры, гранулоциты, тучные и дендритные клетки.
Естественные или нормальные киллеры, NK-клетки (от англ. Naturalkiller) были изначально описаны как большие гранулярные лимфоциты, способные распознать в организме некоторые виды раково-трансформированных клеток и уничтожить их без предварительной подготовки, что обусловило название клеток.
Сейчас установлено, что естественные киллеры (ЕК) имеют морфологию малых лимфоцитов, на долю которых приходится около 15% всех лимфоцитов. Они образуются из ППСК костного мозга, мигрируют с кровотоком, но отсутствуют в лимфе. Обнаруживаются в печени, селезенке, слизистых оболочках, матке. По маркерам, местам типичной локализации и эффекторным механизмам выделяют две субпопуляции ЕК: кровяную и тканевую. ЕК - главный защитник макроорганизма от внутриклеточных паразитов. Он срабатывает задолго до активации адаптивного иммунитета. Вместе с тем биологические возможности ЕК весьма ограничены.
Кровяные ЕК - это активно циркулирующие в кровотоке клетки. Обнаруживаются в красной пульпе селезенки. Несут на себе маркер CD16+CD56мало, FcR к иммуноглобулину класса G, связанному в иммунный комплекс, и рецептор к MHC I класса. При активации синтезируют, накапливая в гранулах, перфорин, гранзимы и гранулизин. Эффекторная функция кровяных ЕК в отношении меченных иммуноглобулинами клеток реализуется в АЗКЦТ (см. раздел 11.3.1). Мишенями являются клетки, инфицированные внутриклеточными паразитами (бактерии, вирусы, простейшие), аллогенные клетки трансплантата и некоторые опухоли.
Рецептор к MHC I класса анализирует плотность его экспрессии на мембране клетки. Дефицит этих молекул, наблюдающийся при раковой трансфорации клеток, также потенцирует цитотоксичность ЕК.
Тканевые ЕК ведут более оседлый образ жизни и обнаруживаются в большом количестве в печени и децидаульной оболочке беременной матки. Несут маркер CD16-CD56много и много Fas -лиганда. Реализуют АНКЦТ (см. раздел 11.3.2). Клетками-мишенями являются лимфоциты, активированные, например, пищевыми антигенами или аллоантигенами плода и экспрессирующие Fas.
Помимо цитотоксических функций, ЕК вырабатывают цитокины (ИЛ-5, 8, γ-ИФН, ФНО, гранулоцит-моноцит-колониестимулирующий фактор-ГМ-КСФ и др.), активирует макрофагально-фагоцитарное звено, развитие иммунного ответа и иммунного воспаления. Эффекторная функция ЕК усиливается цитокинами (ИЛ-2, 4, 10, 12, γ-ИФН и др.).
Фагоциты (см. раздел 9.2.3.1) - самая многочисленная морфологически гетерогенная фракция иммунокомпетентных клеток. Выполняют регуляторную и эффекторную функции. Вырабатывают иммуноцитокины, ферменты, ион-радикалы и другие биологически активные вещества, осуществляют вне- и внутриклеточный киллинг и фагоцитоз. Кроме того, макрофаги являются АПК - обеспечивают процессинг и презентацию антигена Т-хелперам.
Эозинофилы - гранулярные лейкоциты крови. Содержатся в крови, рыхлой соединительной ткани, в большом количестве накапливаются в очагах местного воспаления, вызванного гельминтами, и обеспечивают АЗКЦТ.
Эозинофилы несут на мембране низкоаффинные FcR к IgA или IgE, распознающие паразитов, отмеченных такими антителами. Активированная клетка выделяет ферменты (эозинофильная пероксидаза и коллагеназа) и белковые токсины (главный щелочной протеин, эозинофильный катионный белок и нейротоксин), губительно действующие на гельминты.
Эозинофилы также синтезируют цитокины (ИЛ-3, 5, 8, ГМ-КСФ и др.), стимулирующие клеточное звено иммунитета и образование Т2-хелпера, и липидные медиаторы (лейкотриены, тромбоцитактивирующий фактор и др.), запускающие воспалительную реакцию в месте внедрения гельминта.
Тучные клетки - немигрирующие морфологические элементы неясного происхождения, располагаются оседло вдоль барьерных тканей (laminapropria слизистых оболочек, в подкожной соединительной ткани) и в соединительной ткани кровеносных сосудов. По набору синтезируемых биологически активных соединений и локализации выделяют две разновидности тучных