
Распространены повсеместно, как сапротрофы в почве, на древесине, на разных растительных субстратах. Занимают некоторые ниши, недоступные другим грибам (кератинофильные грибы живут на жив остатках, содержащих кератин). Участвуют в разложении раст остатков, содержащих целлюлозу. Так же в морях и пресных водах на древесине, паразитируют на растениях, животных, человеке. Одновременно наносят вред сельскому хозяйству, но так же — важные продуценты антибиотиков, витаминов, ферментов, кормового белка и тд.Основной признак — формирование в результате плового процесса сумок или асков — замкнутых одноклеточных образований с аскоспорами (обычно 8). Вегетативное тело — разветвленный гаплоидный мицелий из одно- или многоядерных клеток. Цитокинез одновременно с кариокинезом центрипетально (от стенок к центру), в центре септы остается пора (может даже транспортироваться ядро), поры играют важную роль в транспорте в-в по гифам в зону роста. У низших мицелия нет, тело — почкующаяся клетка, иногда образует псевдомицелий. Настоящие ткани формируются редко. В состав стенки входит хитин (в среднем не больше 25%) и глюканы (отличаются от целлюлозы характером связи между мономерами). Бесполое размножение конидиями, конидиеносцы развиваются одиночно, соедниняются в пучки и подушечки. Конидиальное спороношение служит для массового расселения грибов. Типичный половой процесс — гаметангиогамия, но гаметангии аскомицетов не гомологичны настоящим гаметангиям, вероятно происходят от недифференцированных копулирующих ветвей мицелия соматогамных грибов.У низших грибов половой процесс сходен с зигогамией зигомицетов. У высших аскомицетов- женский гаметангий из аскогона и нитевидной трихогины, мужской гаметангий антеридий — одноклеточный. Содержимое антеридия переходит в аскогон, но ядра сразу не сливаются, а объединяются попарно, образуя дикарион. Из аскогона вырастают аскогенные гифы, ядра дикариона синхронно делятся и септы делят гифы на двухъядерные клетки. Конечная клетка гифы загибается, в месте перегиба ядра дикариона делятся, одно ядро переходит в кончик, одно в основание, а по центру остается пара ядер разного пола. Два центральных ядра обособляются септами и развиваются в сумку, ядра дикариона сливаются, затем происходит редукционное дление и митоз — образуется 8 гаплоидных ядер,из которых формируются аскоспоры (часть цитопалзмы обособляется вокруг ядер и одевается оболочкой, а часть остается неиспользованной — эпиплазма. Когда споры созревают в эпиплазме гликоген превращается в сахар и с помощью тургорного давления споры выбрасываются из сумки на расстояние до 10 см и более. Затем дикарион восстанавливается в базальной клетке и на одной гифе таким обр формируется много сумок. В итоге цикл развития состоит из длительной гаплофазы, недолгой дикариотической (аскогенные гифы) и очень короткой диплофазы (молодая сумка с диплоидным ядром). У многих антеридий может отсутствовать или не функционировать, тогда его роль играют конидии, вегетативные гифы или специализированные клетки — спермации. Могут быть гомо- и гетероталличные. Сумки делятся на прототуникатные (тонкая оболочка, не участвует в распространении аскоспор) и эутуникатные (плотные оболочки со спец приспособлениями для вскрывания сумки). У некоторых аскоспор есть придатки для более эффективного распространения. У высших аскомицетов сумки развиваются в плодовых телах и аскостромах. Плодовые тела могут быть полностью замкнутые (клейстотеций), полузамкнутые (перитеций) и открытые (апотеций). Аскострома отличается от плодового тела отсутствием собственной оболочки (перидия). Подкласс Hemiascomycetidae — Голосумчатые. Примитивные аскомицеты без плодовых тел, сумки развиваются на мицелии. Порядок Endomycetales — Эндомицетовые. Сумки расположены на мицелии поодиночке, развиваются из зиготы без участия аскогенных гиф, у многих представителей — дрожжей — мицелия нет, клетки размножаются почкованием, а сумки — одиночные клетки. Большинство сапротрофы, обитают на поверхности плодов и вегетативных частей растений, в нектаре, в почве, паразитов мало. Сюда относятся дрожжи (представлены семейством Saccharomycetaceae — возбудители спиртового брожения и продуценты кормового белка). Их отдельные клетки размножаются почкованием или делением, если они не расходятся — образуется псевдомицелий. Половой процесс — копуляция двух вегетативных клеток. Аскоспоры развиваются в сумках, представленных одиночными клетками. Вероятно это вторично упрощенные организмы из-за обитания в жидких сахаристых средах. У некоторых представителей преобладает гаплофаза и диплоидизация перед образованием сумки (Род Schizosaccharomyces), у других после копуляции гамет идет почкование диплоидной клетки, а затем только образование сумки (Saccharomyces cerevisiae или пекарские дрожжи), у третьих преобладает диплофаза и аскоспоры копулируют уже в сумке (Saccharomycodes ludvigii). Дрожжи распространены на разных субстратах, богатых сахарами. При развитии вызывают спиртовое брожение, что используется в изготовлении алкогольных напитков и хлебопечении. Особенно важны представители рода Saccharomyces, Пекарские дрожжи существуют только в культуре — характерна длительная диплоидная фаза, в сумке обычно 4 аскоспоры и вид гетероталличен. Существует множество рас — хлебопекарные, винные, пивные, спиртовые. Так как дрожжи быстро растут и легко культивируются, их часто используют для изучения биохимических процессов, в генетике и др областях биологии.
Подкласс Euascomycetidae — Эуаскомицеты. Сумки образуются в настоящих плодовых телах, только у немногих примитивных сумки формируются группами на мицелии и не окружены перидием. Три типа настоящих плодовых тел: 1)клейстотеций — полностью замкнут, содержит только сумки, расположенные неупорядоченно или в виде пучков или слоя, стерильных элементов (парафиз) нет, аскоспоры освобождаются при разрушении перидия. 2)перитеций — округлые тела с узким отверстием на вершине, со дна и боковых стенок поднимаются сумки, разделенные стерильными нитевидными элементами — парафизами. Сумки активно выбрасывают аскоспоры, поочередно подрастая к отверстию, затем ножка сумки лизируется. Если сумки прототуникатные, то их стенки быстро лизируются и споры лежат в слизи свободно в плектенциме тела. 3)апотеций — блюдцевидные открытые плодовые тела, на верхней стороне слой сумок и парафиз (называется гимением). Под гимениальным слоем тонкий слой переплетеных гиф -гипотеций или субгимениальный слой. Стерильная часть состоит из оболочки апотеция и мякоти. Аскоспоры выбрасываются одновременно всеми сумками в виде взрыва. Является наиболее совершенным плодовым телом, так как способствует наибольшей продукции спор и обеспечивает их активное расселение. Порядок Eurotiales — Эвроциевые. Образуют клейстотеции на мицелии, мелкие, не превышают 1-2мм. Сумки расположены хаотично, прототуникатные с быстро разрушающимися оболочками, внутри 2-8 аскоспор, освобождаются пассивно. Бесполое размножение конидиями, играет важную роль в расселении. Большинство сапротрофы в почвах и чаще всего на растительных остатках, некоторые термофилы и живут в компосте и влажном сене (до 30-60 градусов). Некоторые образуют плесени на пищевых продуктах, синтетических материалах, вызывают коррозии металлов, патогенны для человека и животных, вызывая сильные дерматомикозы и глубокие микозы. Особенно опасен гистоплазмоз (Emmonsiella capsulata). Важнейшие представители образуют антибиотики, ферменты, органические кислоты и широко используются в промышленности — роды Aspergillus, Penicillium (конидиальные стадии этого типа относят к аскомицетам в случае если у них обнаружены сумчатые стадии, остальные относятся к несовершенным грибам). Emericellopsis terricola — продуцент цефалоспорина С, на основе которого были созданы полусинтетические препараты, способные воздействовать на устойчивые к пенициллину грамотрицательные бактерии и стафилококки. Пеницилл и аспергилл распространены в почве от Арктики до тропиков. Многие представители вызывают порчу продуктов, образуя плесени, некоторые виды патогенны для человека и животных (например A. flavus развивается на плодах арахиса и вызывает поражения печени, обладает канцерогенным свойством).Однако многие широкоиспользуются в микробиологической промышленности, P. chrysogenum продуцирует один из основных антибиотиков, применяемых в медицине. 
Для аспергилла хараткерны простые конидиеносцы, вздутые на вершине в виде пузыря, на которых расположены фиалиды (обр цепочки одноклеточных конидий). Клейстотеции с однослойным или многослойным перидием, желтые, образуются желтоватые или красно-желтые плесени. Аскоспоры от бесцветных до красных или фиолетовых. У пеницилла конидиеносцы в виде кисточки, наиболее простой вид — мутовка фиалид на вершине конидиеносца. В более сложном случае на вершине конидиеносца расположены удлиненные клетки метулы, а на каждой — мутовка фиалид. Клейстотеции желтые, обладают неограниченным ростом. Порядок Erysiphales — Эризифовые или мучнисторосяные.
 Образуют клейстотеции, в которых сумки расположены правильным пучком или слоем. Все — облигатные паразиты высших растений, вызывающие «мучнистую росу». Мицелий находится на поверхности растения, проникая гаусториями в клетки эпидермиса пораженного растения (некоторые внедляются даже в мезофилл). Виды, распространенные в засушливых районах развиваются внутри тканей хозяина — приспособление к низкой влажности. На мицелии — неразветвленные конидиеносцы с цепочками конидий (могут быть одиночные конидии) — они и образуют мучнистый налет. Конидии переносятся воздухом к здоровым растениям, могут развиваться и прорастать в сухую погоду (ксерофилы). В конце периода вегетации на мицелии образуются аскогоны без трихогины и антеридии. Содержимое антеридия переходит в аскогон, который после этого делится на несколько клеток, одна из них содержит дикарион. Клетка с дикарионом либо сама становится сумкой, либо дает аскогенные гифы, на которых развиваются сумки, одновременно формируется перидий. Верхний слой перидия из толстых гиф — защитная функция, внутренний из тонких — питательная функция. Формы придатков на внешнем слое и количество сумок в клейстотеции — родовой признак. Клейстотеций — зимующая стадия, но у некоторых мицелий сохраняется в зимующих частях растения. Аскоспоры выходят активно, сумки набухают и разрывают перидий, а потом и сами взрываются, разбрасывая аскоспоры. До 90% развиваются на двудольных. Род Sphaerotheca — в клейстотеции одна сумка и он имеет придатки, похожие на вегетативные гифы. Наиболее распространен S. mors uvae — поражает крыжовник (ягоды, стебли и листья), покрывая ягоды налетом мицелия. Род Microsphaera — жесткие дихотомически ветвящиеся придатки клейстотеция, в котором несколько сумок. M. alphitoides — возбудитель мучнистой росы дуба, поражает стебли и листья. Род Uncinula — жесткие придатки, спирально закрученные на концах. U. necator — возбудитель пепелицы винограда, приносит большие потери урожая. Так же, как и возбудитель болезни крыжовника был завезен в Европу из Северной Америки. Порядок Clavicipitales — Спорыньевые. Образуют перитеции в развитых стромах, состоящих из гиф гриба, окрашены светло или ярко, имеют разную форму. Стромы развиваются на пораженных органах растения-хозяина или из склероциев. Перитеции типичного строения, светлые или белые, обычно погружены в строму, а на поверхности выступают только носики (могут подниматься над стромой более значительно). Парафизы быстро лизируются, сумки цилиндрические с утолщением оболочки на вершине, аскоспоры всегда нитевидные и с большим количеством поперечных перегородок, после высвобождения распадаются на несколько клеток, каждая может прорасти. Аскоспоры выходят поочередно через отверстие в сумке. Все изученные виды гомоталличны. Большинство — паразиты цветковых растений, грибов и членистоногих, особенно важны паразиты однодольных из семейств злаков и осоковых. Наблюдается строгая органотропная специализаия. Род Claviceps (спорынья) — образует твердые темные склероции в завязи растения, из которых после зимовки развиваются желтые или красноватые головчатые стромы с перитециями. По периферии стромы закладываются камеры, в каждой — многоядерный аскогон без трихогины и на той же гифе многоядерные антеридии. После плазмогамии развиваются аскогенные гифы и сумки, одновременно формируется перидий и отделяет перитеции от ткани стромы. Большинство растут на злаках, особенно C. Рurpurea — поражает пшеницу, рожь, ячмень, пырей итд.
Образуют клейстотеции, в которых сумки расположены правильным пучком или слоем. Все — облигатные паразиты высших растений, вызывающие «мучнистую росу». Мицелий находится на поверхности растения, проникая гаусториями в клетки эпидермиса пораженного растения (некоторые внедляются даже в мезофилл). Виды, распространенные в засушливых районах развиваются внутри тканей хозяина — приспособление к низкой влажности. На мицелии — неразветвленные конидиеносцы с цепочками конидий (могут быть одиночные конидии) — они и образуют мучнистый налет. Конидии переносятся воздухом к здоровым растениям, могут развиваться и прорастать в сухую погоду (ксерофилы). В конце периода вегетации на мицелии образуются аскогоны без трихогины и антеридии. Содержимое антеридия переходит в аскогон, который после этого делится на несколько клеток, одна из них содержит дикарион. Клетка с дикарионом либо сама становится сумкой, либо дает аскогенные гифы, на которых развиваются сумки, одновременно формируется перидий. Верхний слой перидия из толстых гиф — защитная функция, внутренний из тонких — питательная функция. Формы придатков на внешнем слое и количество сумок в клейстотеции — родовой признак. Клейстотеций — зимующая стадия, но у некоторых мицелий сохраняется в зимующих частях растения. Аскоспоры выходят активно, сумки набухают и разрывают перидий, а потом и сами взрываются, разбрасывая аскоспоры. До 90% развиваются на двудольных. Род Sphaerotheca — в клейстотеции одна сумка и он имеет придатки, похожие на вегетативные гифы. Наиболее распространен S. mors uvae — поражает крыжовник (ягоды, стебли и листья), покрывая ягоды налетом мицелия. Род Microsphaera — жесткие дихотомически ветвящиеся придатки клейстотеция, в котором несколько сумок. M. alphitoides — возбудитель мучнистой росы дуба, поражает стебли и листья. Род Uncinula — жесткие придатки, спирально закрученные на концах. U. necator — возбудитель пепелицы винограда, приносит большие потери урожая. Так же, как и возбудитель болезни крыжовника был завезен в Европу из Северной Америки. Порядок Clavicipitales — Спорыньевые. Образуют перитеции в развитых стромах, состоящих из гиф гриба, окрашены светло или ярко, имеют разную форму. Стромы развиваются на пораженных органах растения-хозяина или из склероциев. Перитеции типичного строения, светлые или белые, обычно погружены в строму, а на поверхности выступают только носики (могут подниматься над стромой более значительно). Парафизы быстро лизируются, сумки цилиндрические с утолщением оболочки на вершине, аскоспоры всегда нитевидные и с большим количеством поперечных перегородок, после высвобождения распадаются на несколько клеток, каждая может прорасти. Аскоспоры выходят поочередно через отверстие в сумке. Все изученные виды гомоталличны. Большинство — паразиты цветковых растений, грибов и членистоногих, особенно важны паразиты однодольных из семейств злаков и осоковых. Наблюдается строгая органотропная специализаия. Род Claviceps (спорынья) — образует твердые темные склероции в завязи растения, из которых после зимовки развиваются желтые или красноватые головчатые стромы с перитециями. По периферии стромы закладываются камеры, в каждой — многоядерный аскогон без трихогины и на той же гифе многоядерные антеридии. После плазмогамии развиваются аскогенные гифы и сумки, одновременно формируется перидий и отделяет перитеции от ткани стромы. Большинство растут на злаках, особенно C. Рurpurea — поражает пшеницу, рожь, ячмень, пырей итд. 
Склероции зимуют в почве, во время цветения хозяина аскоспоры переносятся ветром и заражают их. Заражение возможно только при раскрытых цветковых чешуях, спора попадает на рыльце пестика, прорастает и ростовая трубка достигает завязи. Затем там развивается конидиальная стадия и много мелких конидий, покрытых «медвяной росой» - содержит много сахаров и привлекает насекомых, которые и переносят конидии на др растения. Так же могут переноситься ветром и каплями дождя. Склероции развиваются медленно, обычно ко времени спелости зерна. Заражение опасно тем, что семена зараженного растения содержат токсичные алкалоиды, которые вызывают сокращение гладкой мускулатуры и сосудов, а так же воздействуют на НС. Эти алкалоиды используются в медицине для лечения СС и нервных заболеваний. Род Epichloe — развиваются на определенных частях растений — побегах с зачатками соцветий. Наиболее распространена Epichloe typhina — рогозовидная эпихлое, вызывает чехловидную болезнь многолетних злаков. Стромы расположены на стебле и окружают его в виде чехла. На поверхности мелкие одиночные конидии, а в конце лета в строме развиваются перитеции. Мицелий распространяется по межклетникам, а в зоне зачатка соцветия гифы внедряются в клетки сосудов. Порядок Pezizales — Пецициевые. Сумки открываются крышечкой, плодовые тела - апотеции типичного строения, реже — несущие гимений на складчатой шляпке на стерильной ножке(моршеллоидный или гельвеллоидный апотеций). Чаще всего апотеций имеет мясистую консистенцию, окраска от ярко оранжевой до черной. В гимении всегда есть парафизы, чаще всего того же размера, что и сумки. Большинство имеют только сумчатую стадию, только у некоторых есть и конидиальная. Большинство сапротрофы — гумусовые, подстилочные, лигнофилы, карбофилы, копрофилы, благоприятна повышенная влажность, многие развиваются весной. Род Peziza — чашевидные апотеции 1-5 см бурого цвета, гладкие или мучнистые. В основном на влажной почве, некоторые лигнофилы — на гнилой древесине. Род Sclerotinia — в процессе развития проходят две стадии: во время первой образует покоящиеся структуры — склероции, во время второй — апотеции в форме бокала или блюдца, где развиваются цилиндрические сумки с аскоспорами. Размножение у большинства видов конидиями. Они поражают, в частности, такие культуры, как подсолнечник, картофель, капуста, морковь, фасоль, горох, плоды фруктовых растений (виноград). Склеротиния также поражает и другие культурные и дикорастущие растения (например брусника, клюква, черника, берёза). Род  Morchella (сморчок) — крупные складчатые апотеции на стерильной ножке, шляпка яйцевидная, окраска от желто-бурой до бурой, складки выстланы гимением, а разделяющие ребра остаются стерильными. Выбрасывание аскоспор постепенное. Наиболее распространенные виды — M. esculenta и M. conica. Первый встречается весной под лиственными деревьями, второй — в смешанных или хвойных лесах на только что прогретой земле. Все сморчки съедобны. Род Gyromitra (строчок) — крупные апотеции неправильной формы, шляпка с неупорядоченной складчатостью, бурого цвета, ножка толстая и бороздчатая. Сапротрофы на почве, развиваются весной. Наиболее распространен вид G. esculenta. Считается условно съедобным грибом из-за различного содержания токсина гиромитрина. Род Tuber - Плодовые тела трюфелей располагаются под землёй, имеют округлую или клубневидную форму и мясистую консистенцию. Апотеции в зрелом состоянии у большинства видов замкнуты. Наружная часть— перидий, который может быть снаружи гладким, растрескивающимся или же покрытым более или менее крупными полиэдрическими бородавками. Ткань плодового тела состоит из чередующихся светлых и тёмных прожилок; светлые прожилки называют «внутренними венами», а тёмные — «наружными венами». Сумки располагаются на внутренних венах и имеют разнообразную форму. Растут в лиственных лесах и образуют микоризу с корнями деревьев.
Morchella (сморчок) — крупные складчатые апотеции на стерильной ножке, шляпка яйцевидная, окраска от желто-бурой до бурой, складки выстланы гимением, а разделяющие ребра остаются стерильными. Выбрасывание аскоспор постепенное. Наиболее распространенные виды — M. esculenta и M. conica. Первый встречается весной под лиственными деревьями, второй — в смешанных или хвойных лесах на только что прогретой земле. Все сморчки съедобны. Род Gyromitra (строчок) — крупные апотеции неправильной формы, шляпка с неупорядоченной складчатостью, бурого цвета, ножка толстая и бороздчатая. Сапротрофы на почве, развиваются весной. Наиболее распространен вид G. esculenta. Считается условно съедобным грибом из-за различного содержания токсина гиромитрина. Род Tuber - Плодовые тела трюфелей располагаются под землёй, имеют округлую или клубневидную форму и мясистую консистенцию. Апотеции в зрелом состоянии у большинства видов замкнуты. Наружная часть— перидий, который может быть снаружи гладким, растрескивающимся или же покрытым более или менее крупными полиэдрическими бородавками. Ткань плодового тела состоит из чередующихся светлых и тёмных прожилок; светлые прожилки называют «внутренними венами», а тёмные — «наружными венами». Сумки располагаются на внутренних венах и имеют разнообразную форму. Растут в лиственных лесах и образуют микоризу с корнями деревьев.
Класс Basidiomycetes — Базидиомицеты. Высшие грибы с многоклеточным мицелием. Половое спороношение — базидиоспорами (экзогенные споры, образующиеся на специальных выростах мицелия базидиях). Половых органов нет, сливаются две вегетативные клетки гаплоидного мицелия (из базидиоспор). Большинство гетероталличны, сливаются только клетки гиф из базидиоспор разных половых знаков (+ и -). Сливается только цитоплазма, ядра объединяются в дикарионы и в таком состоянии мицелий может находиться длительное время (дикарионы синхронно делятся). Базидии образуются из двухъядерных клеток на концах гиф (напоминают сумку аскомицетов и гомологичны ей). У апикальной двухъядерной клетки гифы образуется вырост (пряжка)(А, Б), одновременно делятся ядра дикариона и сама клетка делится септой — в итоге в верхней клетке остается два не сестринских ядра, одно ядро 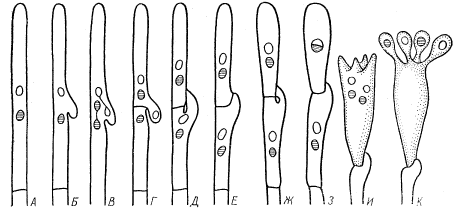 переходит в пряжку, а последнее ядро оказывается отделено септой (В, Г). Затем пряжка отделяется септой от апикальной клетки и сливается с нижележащей клеткой, восстанавливая ее двухъядерность, чтобы та могла образовать новые базидии. В то же время в апикальной клетке ядра сливаются, делятся редукционно, клетка вытягивается и становится базидией — гаплоидные ядра переходят в выросты (стеригмы) на вершине (развиваются в базидиоспоры) (И, К). При созревании базидиоспоры отбрасываются на небольшое расстояние.
переходит в пряжку, а последнее ядро оказывается отделено септой (В, Г). Затем пряжка отделяется септой от апикальной клетки и сливается с нижележащей клеткой, восстанавливая ее двухъядерность, чтобы та могла образовать новые базидии. В то же время в апикальной клетке ядра сливаются, делятся редукционно, клетка вытягивается и становится базидией — гаплоидные ядра переходят в выросты (стеригмы) на вершине (развиваются в базидиоспоры) (И, К). При созревании базидиоспоры отбрасываются на небольшое расстояние.  По строению три типа базидий: 1) холобазидия — одноклеточная цилиндрическая, споры расположены на одном уровне (акроспоровая базидия) или без особого порядка на боковых сторонах (плевроспоровая базидия). 2) гетеробазидия — состоит из гипобазидии и эпибазидии (из двух — четырех частей), которые могут быть разделены перегородкой. 3)телиобазидия или фрагмобазидия — разделена поперечными перегородками на четыре клетки, по бокам которых формируются споры. Обычно образуется из толстостенной покоящейся клетки телиоспоры (поэтому называют телиобазидией). Чаще всего базидии возникают на или внутри плодовых тел, сложенных из дикариотичного мицелия. Таким образом преобладает дикариотичная фаза, гаплоидная — только базидиоспора и мицелий, вырастающий из нее на короткий срок. Плодовые тела очень разнообразны. Спороносный слой — гимений расположен либо на верхней (примитивный признак) либо на нижней стороне тела. Гимений состоит из базидий со спорами, базидиол (молодые базидии) и парафиз, разделяющих базидии друг от друга. У некоторых есть цистиды — крупные клетки, возвышаются над гимениальным слоем и защищают базидии. Поверхность, несущая гимений — гименофор. У примитивных видов он гладкий, но эволюция шла по пути увеличения площади гименофора за счет образования складок и шипов, а так же по пути перемещения гименофора с верхней поверхности на нижнюю. Активное отбрасывание спор, как и у аскомицетов, за счет гидролиза гликогена и повышения тургорного давления, но так как канал стеригмы очень узкий — получается небольшой толчок. В закрытых плодовых телах правильный гимений часто не образуется. В зависимости от цвета зрелых базидиоспор гименофор может быть различно окрашен (от бесцветных до темно-фиолетовых и черных). Конидиальное спороношение довольно редко, могут развиваться и на гаплоидных, и на диплоидных мицелиях. Главные отличия от аскомицетов: 1)у базидиомицетов половое спороношение экзогенное, а у аскомицетов — эндогенное. 2) у аскомицетов встречаются половые органы, у базидиомицетов - соматогамия. 3)У аскомицетов преобладает гаплоидная стадия, у базидиомицетов — дикариотичная. 4) плодовые тела аскомицетов из гаплоидного мицелия, дикариточны только аскогенные гифы, а у базидиомицетов плодовые тела полностью образованы дикариотичными гифами. Классификация по типу развития и строения базидии. Базидиомицеты в основном сапротрофы, играют важную роль в минерализации органических соединений, в особенности трудноразрушимых (целлюлоза, лигнин). Грибы выделяют свои ферменты в среду и впитывают всей поверхностью продукты распада органики, разрушая при этом гораздо больше вещества чем реально используют. Наибольшее распространение базидиомицеты имеют в лесных почвах и подстилке. Многие базидиомицеты выступают в качестве симбионтов растений, другие — паразитов растений (опёнок, трутовик, головня, ржавчинные грибы). Подкласс Holobasidiomycetidae — Холобазидиомицеты. Грибы с неразделенной одноклеточной булавовидной или цилиндрической базидией, классификация основана на расположении базидий. Группа порядков Гименомицеты. Самая большая группа, то, что называем грибами — плодовые тела, а сама грибница в субстрате — почве, древесине итд. Гименофор очень разнообразен от гладкого до образующего различные складки, трубочки и пластинки. Плодовые тела в виде распростертых по субстрату корочек (ресупинатные), прямостоячие разветвленные, деревянистые копытообразные (на стволах деревьев), очень много видов с плодовыми телами мясистой консистенции и состоящих из шляпки и ножки. Эволюция шла от коркообразных тел с гименофором на всей поверхности к шляпочным телам, которые возвышаются над субстратом на ножке, а гименофор формируется на нижней стороне шляпки, что защищает его и облегчается распространение базидиоспор. Гимений из базидий, базидиол, парафиз и часто цистид. Гимений либо с самого начала лежит открыто, либо в молодом возрасте прикрыт сплетением гиф — покрывалом, но к моменту созревания всегда открывается. Большая часть плодового тела состоит из стерильных гиф, поверхностные могут иметь окрашенные оболочки. Тела могут быть однолетними и многолетними (у трутовиков до 80 лет). Очень характерно образование гигантских плодовых тел. Окраска базидиоспор (и соотв гименофора) белая, розовая, коричневая, фиолетово-черная. Споры разносятся ветром, а у мясистых плодов еще и благодаря животным, поедающим их. Гименомицеты очень распространены в лесу, на лугах, в степях, даже полупустынях и путсынях, большинство сапротрофы на мертвой древесине, листовом опаде и в почве, разлагают органические остатки. Некоторые паразиты, поселяются на корнях и стволах живых деревьев (корневая губка, опенок осенний). Очень многие образуют микоризу: 1) эндотрофная — мицелий в основном в паренхиме корня растения, при этом корневые волоски сохраняются. 2) эктотрофная — на поверхности корня образуется чехол из гиф, волоски при этом не образуются, а гифы гриба походят между клетками эидермиса. 3) экто-эндотрофная — промежуточный тип, встречается чаще остальных. В зависимости от субстрата различают ксилофилов (живут на древесине), почвенных сапротрофов, микоризные грибы, копрофилов (на навозе животных), карбофилов (на обугленной древесине), микофилов (на других гименомицетах). Значение — съедобные плодовые тела (шампиньоны, белые, подберезовики, вешенка и тд), некоторые продуцируют целюлолитические ферменты и др биоактивные в-ва. Так же могут наносить вред, разрушая деревянные постройки. Преобладают гетероталличные виды. Порядок Aphyllophorales — Афиллофоровые.
По строению три типа базидий: 1) холобазидия — одноклеточная цилиндрическая, споры расположены на одном уровне (акроспоровая базидия) или без особого порядка на боковых сторонах (плевроспоровая базидия). 2) гетеробазидия — состоит из гипобазидии и эпибазидии (из двух — четырех частей), которые могут быть разделены перегородкой. 3)телиобазидия или фрагмобазидия — разделена поперечными перегородками на четыре клетки, по бокам которых формируются споры. Обычно образуется из толстостенной покоящейся клетки телиоспоры (поэтому называют телиобазидией). Чаще всего базидии возникают на или внутри плодовых тел, сложенных из дикариотичного мицелия. Таким образом преобладает дикариотичная фаза, гаплоидная — только базидиоспора и мицелий, вырастающий из нее на короткий срок. Плодовые тела очень разнообразны. Спороносный слой — гимений расположен либо на верхней (примитивный признак) либо на нижней стороне тела. Гимений состоит из базидий со спорами, базидиол (молодые базидии) и парафиз, разделяющих базидии друг от друга. У некоторых есть цистиды — крупные клетки, возвышаются над гимениальным слоем и защищают базидии. Поверхность, несущая гимений — гименофор. У примитивных видов он гладкий, но эволюция шла по пути увеличения площади гименофора за счет образования складок и шипов, а так же по пути перемещения гименофора с верхней поверхности на нижнюю. Активное отбрасывание спор, как и у аскомицетов, за счет гидролиза гликогена и повышения тургорного давления, но так как канал стеригмы очень узкий — получается небольшой толчок. В закрытых плодовых телах правильный гимений часто не образуется. В зависимости от цвета зрелых базидиоспор гименофор может быть различно окрашен (от бесцветных до темно-фиолетовых и черных). Конидиальное спороношение довольно редко, могут развиваться и на гаплоидных, и на диплоидных мицелиях. Главные отличия от аскомицетов: 1)у базидиомицетов половое спороношение экзогенное, а у аскомицетов — эндогенное. 2) у аскомицетов встречаются половые органы, у базидиомицетов - соматогамия. 3)У аскомицетов преобладает гаплоидная стадия, у базидиомицетов — дикариотичная. 4) плодовые тела аскомицетов из гаплоидного мицелия, дикариточны только аскогенные гифы, а у базидиомицетов плодовые тела полностью образованы дикариотичными гифами. Классификация по типу развития и строения базидии. Базидиомицеты в основном сапротрофы, играют важную роль в минерализации органических соединений, в особенности трудноразрушимых (целлюлоза, лигнин). Грибы выделяют свои ферменты в среду и впитывают всей поверхностью продукты распада органики, разрушая при этом гораздо больше вещества чем реально используют. Наибольшее распространение базидиомицеты имеют в лесных почвах и подстилке. Многие базидиомицеты выступают в качестве симбионтов растений, другие — паразитов растений (опёнок, трутовик, головня, ржавчинные грибы). Подкласс Holobasidiomycetidae — Холобазидиомицеты. Грибы с неразделенной одноклеточной булавовидной или цилиндрической базидией, классификация основана на расположении базидий. Группа порядков Гименомицеты. Самая большая группа, то, что называем грибами — плодовые тела, а сама грибница в субстрате — почве, древесине итд. Гименофор очень разнообразен от гладкого до образующего различные складки, трубочки и пластинки. Плодовые тела в виде распростертых по субстрату корочек (ресупинатные), прямостоячие разветвленные, деревянистые копытообразные (на стволах деревьев), очень много видов с плодовыми телами мясистой консистенции и состоящих из шляпки и ножки. Эволюция шла от коркообразных тел с гименофором на всей поверхности к шляпочным телам, которые возвышаются над субстратом на ножке, а гименофор формируется на нижней стороне шляпки, что защищает его и облегчается распространение базидиоспор. Гимений из базидий, базидиол, парафиз и часто цистид. Гимений либо с самого начала лежит открыто, либо в молодом возрасте прикрыт сплетением гиф — покрывалом, но к моменту созревания всегда открывается. Большая часть плодового тела состоит из стерильных гиф, поверхностные могут иметь окрашенные оболочки. Тела могут быть однолетними и многолетними (у трутовиков до 80 лет). Очень характерно образование гигантских плодовых тел. Окраска базидиоспор (и соотв гименофора) белая, розовая, коричневая, фиолетово-черная. Споры разносятся ветром, а у мясистых плодов еще и благодаря животным, поедающим их. Гименомицеты очень распространены в лесу, на лугах, в степях, даже полупустынях и путсынях, большинство сапротрофы на мертвой древесине, листовом опаде и в почве, разлагают органические остатки. Некоторые паразиты, поселяются на корнях и стволах живых деревьев (корневая губка, опенок осенний). Очень многие образуют микоризу: 1) эндотрофная — мицелий в основном в паренхиме корня растения, при этом корневые волоски сохраняются. 2) эктотрофная — на поверхности корня образуется чехол из гиф, волоски при этом не образуются, а гифы гриба походят между клетками эидермиса. 3) экто-эндотрофная — промежуточный тип, встречается чаще остальных. В зависимости от субстрата различают ксилофилов (живут на древесине), почвенных сапротрофов, микоризные грибы, копрофилов (на навозе животных), карбофилов (на обугленной древесине), микофилов (на других гименомицетах). Значение — съедобные плодовые тела (шампиньоны, белые, подберезовики, вешенка и тд), некоторые продуцируют целюлолитические ферменты и др биоактивные в-ва. Так же могут наносить вред, разрушая деревянные постройки. Преобладают гетероталличные виды. Порядок Aphyllophorales — Афиллофоровые.  Плодовые тела разнообразны, гименофор гладкий, бугорчатый, складчатый, трубчатый, реже плстинчатый.Кроме базидий, базидиол и парафиз у некоторых есть цистиды, щетинки и другие элементы защиты. Гифы тонкие, часто бесцветные. Морфологически плодовые тела бывают распростертые, чашевидные с гименофором на внутренней плоскости, шляповидные с зачаточной боковой ножкой или центральной ножкой и гименофором на нижней поверхности, прямостоячие цилиндрические или разветвленные с гименофором на верхней части тела. Большинство сапротрофы, микоризообразователи, часть — паразиты деревьев и травянистых растений. Распространены во всех природных зонах, но особенно широко в лесах, где являются основными разрушителями древесины. В основе классификации — строение и расположение гименофора, консистенция плодовых тел, окраска спор, онтогенез. Трутовики (Семейство Polyporaceae) — сапротрофы на мертвой древесине и паразиты деревьев. Плодовые тела почковидные, шляпковидные, копытообразные, гименофор в виде радиальных или переплетающихся складок, ячеек, трубочек. Способны активно разрушать древесину с помощью экзоферментов, делятся на целлюлозоразрушающую и лигнинразрушающую группу, вызывают разные типы гнили. Мицелий развивается внутри субстрата, плодовые тела образуются на поверхност, ориентированы гименофором вниз. Тело может быть однолетним и многолетним. У многолетних плодовых тел с трубчатым гименофором прошлогодний слой трубочек каждый год зарастает мицелием, уплотняется и образуется новый слой, который начинает функционировать в середине лета. У плодовых тел со складчатым гименофором слой один, но каждый год он удлинняется. Род Fomitopsis – трубчатый гименофор, нарастающий каждый год, ткань окрашена в светлые тона. F. рinicola трутовик окаймленный почти черное плодовое тело с яркой оранжевой каймой по краю. Встречается на хвойных и лиственных породах в лесах умеренной зоны, растет на мертвой древесине или празитирует на ослабленных живых деревьях. Род Fomes — рыжеватое плодовое тело. F. fomentarius — трутовик настоящий, преимущественно на ослабленных или мертвых березах. На поверхности заметны зоны ежегодного прироста гименофора. Домовый гриб (Serpula lacrymans) — весьма вредоносный гриб-дереворазрушитель. Поселяется на мёртвом (срубленном) дереве, особенно опасен при появлении в постройках. Чаще всего гриб развивается в тёмных, душных и влажных погребах и подвалах, у основания балок, на нижней поверхности досок пола, непосредственно покоящегося на влажной почве. Для истребления уже появившегося домового гриба можно пропитывать куски дерева креозотом или так называемым карболинеумом (Carbolineum), обмазывать обыкновенным дёгтем; другие исследователи указывают на петролеум, как на хорошее средство. Пока гриб не особенно распространился, с успехом можно применить тщательное вырезание повреждённых кусков и замену их новыми. В начале развития гриба на дереве появляются белые маленькие точки, которые со временем сливаются в слизистые пятна или шерстистые нежные налеты, затем появляется сплетение, похожее на паутину серебристого цвета. Постепенно оно все больше и больше распространяется по поверхности дерева, утолщается, приобретает листоватую структуру, шелковистый блеск и пепельно-серый цвет. От краёв гриба отходят тонкие нити, переползающие в поисках за пищей через мельчайшие отверстия и трещины в каменных стенах из одной части дома в другую. Порядок Agaricales — Агариковые.
Плодовые тела разнообразны, гименофор гладкий, бугорчатый, складчатый, трубчатый, реже плстинчатый.Кроме базидий, базидиол и парафиз у некоторых есть цистиды, щетинки и другие элементы защиты. Гифы тонкие, часто бесцветные. Морфологически плодовые тела бывают распростертые, чашевидные с гименофором на внутренней плоскости, шляповидные с зачаточной боковой ножкой или центральной ножкой и гименофором на нижней поверхности, прямостоячие цилиндрические или разветвленные с гименофором на верхней части тела. Большинство сапротрофы, микоризообразователи, часть — паразиты деревьев и травянистых растений. Распространены во всех природных зонах, но особенно широко в лесах, где являются основными разрушителями древесины. В основе классификации — строение и расположение гименофора, консистенция плодовых тел, окраска спор, онтогенез. Трутовики (Семейство Polyporaceae) — сапротрофы на мертвой древесине и паразиты деревьев. Плодовые тела почковидные, шляпковидные, копытообразные, гименофор в виде радиальных или переплетающихся складок, ячеек, трубочек. Способны активно разрушать древесину с помощью экзоферментов, делятся на целлюлозоразрушающую и лигнинразрушающую группу, вызывают разные типы гнили. Мицелий развивается внутри субстрата, плодовые тела образуются на поверхност, ориентированы гименофором вниз. Тело может быть однолетним и многолетним. У многолетних плодовых тел с трубчатым гименофором прошлогодний слой трубочек каждый год зарастает мицелием, уплотняется и образуется новый слой, который начинает функционировать в середине лета. У плодовых тел со складчатым гименофором слой один, но каждый год он удлинняется. Род Fomitopsis – трубчатый гименофор, нарастающий каждый год, ткань окрашена в светлые тона. F. рinicola трутовик окаймленный почти черное плодовое тело с яркой оранжевой каймой по краю. Встречается на хвойных и лиственных породах в лесах умеренной зоны, растет на мертвой древесине или празитирует на ослабленных живых деревьях. Род Fomes — рыжеватое плодовое тело. F. fomentarius — трутовик настоящий, преимущественно на ослабленных или мертвых березах. На поверхности заметны зоны ежегодного прироста гименофора. Домовый гриб (Serpula lacrymans) — весьма вредоносный гриб-дереворазрушитель. Поселяется на мёртвом (срубленном) дереве, особенно опасен при появлении в постройках. Чаще всего гриб развивается в тёмных, душных и влажных погребах и подвалах, у основания балок, на нижней поверхности досок пола, непосредственно покоящегося на влажной почве. Для истребления уже появившегося домового гриба можно пропитывать куски дерева креозотом или так называемым карболинеумом (Carbolineum), обмазывать обыкновенным дёгтем; другие исследователи указывают на петролеум, как на хорошее средство. Пока гриб не особенно распространился, с успехом можно применить тщательное вырезание повреждённых кусков и замену их новыми. В начале развития гриба на дереве появляются белые маленькие точки, которые со временем сливаются в слизистые пятна или шерстистые нежные налеты, затем появляется сплетение, похожее на паутину серебристого цвета. Постепенно оно все больше и больше распространяется по поверхности дерева, утолщается, приобретает листоватую структуру, шелковистый блеск и пепельно-серый цвет. От краёв гриба отходят тонкие нити, переползающие в поисках за пищей через мельчайшие отверстия и трещины в каменных стенах из одной части дома в другую. Порядок Agaricales — Агариковые.  Плодовые тела однолетние, состоят из шляпки и ножки, мягкие. Сидячие тела без ножки характерны для древесных видов (вешенка). Гименофор на нижней стороне, в виде радиально расположенных пластинок. Каждая пластинка в сечении имеет вид конуса, а с двух сторон — гимении. Центр стерилен и называется трамой, есть четыре типа: 1) неправильная трама — из переплетающихся гиф неправильной формы. 2) правильная трама — гифы с цилиндрическими клетками, направлены параллельно длине пластинок. 3)билатеральная трама — в ценре параллельные гифы, от которых расходятся гифы к бокам. 4) инвертная трама — то же, что билатеральная, но боковые гифы идут не от центра к бокам, а от боков к центру. По расположению относительно ножки пластинки бывают свободными (не достигают ножки), могут прикрепляться к ней, могут быть нисходящими по ножке. Когда гименофор залагается открыто — гимнокарпное плодовое тело, если сначала прикрыт сплетением гиф (покрывалом) — гемиангиокарпное тело. Покрывало бывает общее (покрывает и ножу, и шляпку), по мере роста оно разрывается и остается в виде влагалища у основания ножки. Частное покрывало соединяет края шляпки с ножкой и укрывает только формирующийся гименофор. При разрывании оно остается на ножке в виде кольца или как бахрома по краю шляпки. Агариковые — сапротрофы на почве, опаде, древесине, микоризообразователи, немногие — паразиты. Именно сюда относятся основные шляпочные грибы, съедобные и ядовитые. Съедобные грибы: Семейство болетовые (Boletaceae — трубчатый гименофор, отделяется от мякоти плодового тела, споровый порошок бурый, желтый или розовый) - Белый гриб(Boletus edulis), подосиновик (Leccinum aurantiacum), подберезовик (L. scabrum), виды рода масленок; Семейство свинушковые (Paxillaceae);Семейство шампиньоновые (Agaricaceae — пластинки свободные, всегда есть частное покрывало, окраска спор от белой до черно-бурой): шампиньон двуспоровый (Agaricus bisporus), Agaricus campester; Семейство сыроежковые (Russulaceae): сыроежка, груздь настоящий, рыжик, волнушка. Ядовитые грибы:Семейство мухоморовые (Amanitaceae):бледная поганка, мухомор вонючий, мухомор красный, поганковидный;
Плодовые тела однолетние, состоят из шляпки и ножки, мягкие. Сидячие тела без ножки характерны для древесных видов (вешенка). Гименофор на нижней стороне, в виде радиально расположенных пластинок. Каждая пластинка в сечении имеет вид конуса, а с двух сторон — гимении. Центр стерилен и называется трамой, есть четыре типа: 1) неправильная трама — из переплетающихся гиф неправильной формы. 2) правильная трама — гифы с цилиндрическими клетками, направлены параллельно длине пластинок. 3)билатеральная трама — в ценре параллельные гифы, от которых расходятся гифы к бокам. 4) инвертная трама — то же, что билатеральная, но боковые гифы идут не от центра к бокам, а от боков к центру. По расположению относительно ножки пластинки бывают свободными (не достигают ножки), могут прикрепляться к ней, могут быть нисходящими по ножке. Когда гименофор залагается открыто — гимнокарпное плодовое тело, если сначала прикрыт сплетением гиф (покрывалом) — гемиангиокарпное тело. Покрывало бывает общее (покрывает и ножу, и шляпку), по мере роста оно разрывается и остается в виде влагалища у основания ножки. Частное покрывало соединяет края шляпки с ножкой и укрывает только формирующийся гименофор. При разрывании оно остается на ножке в виде кольца или как бахрома по краю шляпки. Агариковые — сапротрофы на почве, опаде, древесине, микоризообразователи, немногие — паразиты. Именно сюда относятся основные шляпочные грибы, съедобные и ядовитые. Съедобные грибы: Семейство болетовые (Boletaceae — трубчатый гименофор, отделяется от мякоти плодового тела, споровый порошок бурый, желтый или розовый) - Белый гриб(Boletus edulis), подосиновик (Leccinum aurantiacum), подберезовик (L. scabrum), виды рода масленок; Семейство свинушковые (Paxillaceae);Семейство шампиньоновые (Agaricaceae — пластинки свободные, всегда есть частное покрывало, окраска спор от белой до черно-бурой): шампиньон двуспоровый (Agaricus bisporus), Agaricus campester; Семейство сыроежковые (Russulaceae): сыроежка, груздь настоящий, рыжик, волнушка. Ядовитые грибы:Семейство мухоморовые (Amanitaceae):бледная поганка, мухомор вонючий, мухомор красный, поганковидный;
Группа порядков Гастеромицеты.

Плодовые тела полностью замкнуты до созревания спор (ангиокарпные). Гименофор расположен внутри и к моменту созревания спор разрушается, а споры выходят при разрушении или разрыве в оболочке плодового тела. Базидии не участвуют в рассеивании спор, неправильной формы и могут нести больше 4х спор (почти у всех темно окрашенные). Плодовые тела разнообразны по форме, часто в начале развития они шаровидные, но потом меняют свою форму (бокаловидная, блюдцевидная, звездчатая). Диаметр от 2-3 мм до 70 см. Плодовые тела могут быть подземными, полуподземными или наземными. Наземные могут быть сидячими, иметь ложную ножку (стерильный отросток) или настоящую. У некоторых образуется особый плодоносец — рецептакул, несущий спороносную часть плодового тела(часто имеет ячеистую или губчатую структуру). Перидий хорошо развит, может быть одно- или многослойным, различают экзоперидий (гладкий или покрыт бородавочками, шипиками и бугорками) и эндоперидий. Эндоперидий открывается нескоькими способами: правильными или неправильными, одиночными или многочисленными отверстиями, продольными трещинами или экваториальным разрывом (верхняя часть исчезает), часто отверстие на вершине плодового тела снабжено перистомом (вытянутые хоботок). Внутренняя часть плодового тела — глеба — сначала светлая и рыхлая, но по мере созревания базидиоспор темнеет и в ней образуются полости различной формы — камеры. Поверхность выстлана гимением из базидий неправильной формы, которые могут и не образовывать гимения, располагаясь хаотично. Камеры разделены трамами (стерильные участки). При созревании трамы и базидии у большинства разрушаются и базидиоспоры свободно лежат внутри перидия. У некоторых часть трамы идет на формирование капиллиция — особых волокон, разрыхляющих массу спор и участвующих в расселении (при уменьшении влажности нити сокращаются, а при увеличении удлинняются). У некоторых трамы сохраняются, так же иногда глеба распадается на округлые самостоятельные участки, одетые собственной оболочкой — перидиоли (лежат свободно или прикреплены к стенке перидия). Несколько типов развития и расположения спороносного слоя в глебе: 1)равномерный — базидии распределены равномерно и беспорядочно 2)лакунарный — наличие камер или щелей, покрытых базидиями 3) кораллоидный — в центре глебы бесплодная часть, от которой идут коралловидные полости, покрытые гимением 4) многошляпочный — спороносные участки формируются в нескольких местах независимо друг от друга 5)одношляпочный. Эволюция шла по развитию приспособлений для распостранения спор. У видов с подземными плодовыми телами распространение с помощью животных (тело имеет приятный вкус, поедается, споры выходят с экскрементами).У большинства с наземными телами рассеивание с помощью ветра, у степных — отрывается вся верхняя часть и поры сдуваются, а у некоторых плодовые тела отрываются от субстрата и катятся, рассеивая семена. Споры выбрасываются постепенно. Гастеромицеты в основном почвенные сапротрофы, мицелий хорошо развит и пронизывает субстрат, относительно которого встречаются ксилофилы (растительные остатки), почвенные сапротрофы (больше всего), микоризообразователи с древесными породами. В основном сухолюбивые и теплолюбивые, особенно разнообразны в тропических лесах и пустынях. Порядок Lycoperdales- Дождевиковые. Плодовые тела напочвенные, шаровидные, булавовидные звездообразные, сидячие или с ложной ножкой. Перидий двух-четырехслойный, глеба из извилистых камер и трам, при созревании превращаются в порошок. Гимений правильный, хорошо развит капеллиций. Включает роды Lycoperdon (дождевик), Bovista (порховка), Geastrum(звездовик). У звездовика при созревании экзоперидий разрывается на лопасти и отгибается книзу в виде звезды. Молодые плодовые тела употребляются в пищу.
 |
Подкласс Teliobasidiomycetidae — Телиобазидиомицеты. Базидия вырастает из толстостенной покоящейся споры — телиоспоры. Эти споры выполняют функцию покоящихся стадий для перенесения неблагоприятных условий, многие грибы зимуют таким образом. Плодовые тела утрачены в связи с переходом к паразитизму. Телиоспора рассматривается как предшественник базидии — пробазидия. Порядок Ustilaginales — Головневые. Поразиты многих цветковых растений, поражают цветки, семена, листья и стебли, иногда корни. Разрушенные репродуктивные органы превращаются в пылящую массу телиоспор. Мицелий идет по межклетникам, запуская гаустории в клетки. Поначалу заражение может быть незаметно, но затем становится видно, что колоски заполнены не зернами, а черной массой телиоспор. Телиоспоры прорастают четырехклеточной, реже одноклеточной базидией. Род Ustilago — крупные темноокрашенные телиоспоры. Ustilago tritici — возбудитель пыльной головни пшеницы, телиоспоры должны попасть на рыльце цветков, где прорастают четырехклеточной базидией. Базидиоспоры не развиваются, соседние споры или соседние клетки копулируют и сразу образуется дикарион, вырастает дикариотический мицелий и проникает в завязь. При этом семя развивается нормально и внешне выглядит здоровым, но содержит мицелий гриба. Попав в землю, это семя прорастает с виду нормальным растением, однако мицелий проникает в залагающийся колос, разрушает зерна и распадается на телиоспоры. От колоса остается лишь стержень и разрушенные боковые колоски, остальное — телиоспоры. Они переносятся ветром на другие цветущие в это время колосья. Ustilago zeae — возбудитель пузырчатой головни кукурузы, местные поражения разных органов как молодых, так и взрослых растений. Телиоспора прорастает четырехклеточной базидией, базидиоспоры почкуются и эти клетки, опадая, разносятся на разные органы кукурузы, копулируют, имея разные половые знаки. Дикариотичный мицелий поражает ткани, вызывая вздутия, заполненные телиоспорами. Род Tilletia — крупные телиоспоры прорастают одноклеточной базидией с четырьмя базидиеспорами. Tilletia caries — возбудитель твердой головни пшеницы, заражение проростков происходит в почве.
Sphacelotheca panici-miliacei — возбудитель головни проса, телиоспоры прорастают четырехклеточной базидией. Базидиоспоры копулируют, дикариотический мицелий проникает в проросток до его выхода на поверхность почвы. Проникает в конус нарастания и ко времени цветения распадается на телиоспоры. Выходят при выколашивании больного растения.
 |
В следствие параллельной эволюции грибов и питающих растений, у них близкие требования к условиям — температура прорастания, рассеивание телиоспор одновременно с цветением нужных растений и тд. В систему пр