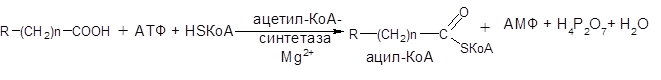1) Образовавшийся оксалоацетат локализован в митохондрии. Мембрана митохондрий непроницаема для образовавшегося оксалоацетата. Последний здесь же в митохондрии восстанавливается в яблочную кислоту:
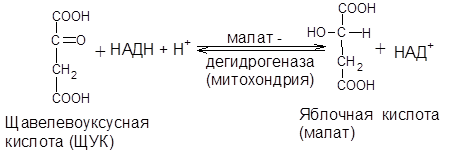
Реакция протекает при участии митохондриальной НАД-зависимой малатдегидрогеназы. Образовавшаяся яблочная кислота легко выходит из митохондрии в цитозоль клетки и вновь окисляется в щавелевоуксусную кислоту при участии цитоплазматической НАД-зависимой малатдегидрогеназы:
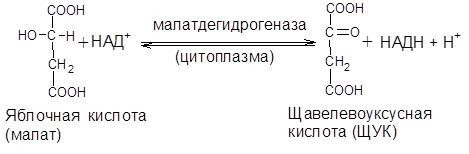
2) Превращение оксалоацетата в фосфоенолпируват происходит в цитоплазме клетки:
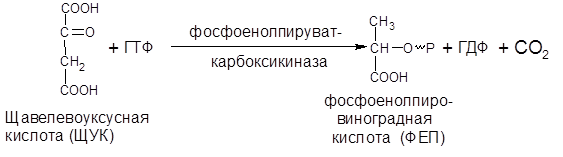
3) Образовавшийся фосфоенолпируват (ФЕП) по пути гликолиза превращается в ПВК, а пируват в результате окислительного декарбоксилирования в ацетил-КоА, который поступая в цикл Кребса, полностью окисляется до СО2 и Н2О, с выделением энергии.
С учетом выше сказанного, видно, что образовавшийся пропионил, в конечном счете, должен превратиться в ацетил-КоА и окислиться в цикле Кребса. Таким образом, все жирные кислоты в результате b-окисления превращаются в ацетил-КоА, который сгорает в цикле Кребса.
Синтез триацилглицеролов
В организме человека исходным сырьем для биосинтеза жиров могут служить углеводы, поступающие с пищей, в растениях – сахароза, поступающая из фотосинтезирующих тканей. Основные этапы синтеза жира включают образование из углеводов глицерол-3-фосфата и жирных кислот, а затем сложноэфирных связей между спиртовыми группами глицерола и карбоксильными группами жирных кислот:

Рисунок 10 – Общая схема синтеза жира из углеводов
Рассмотрим более подробно основные этапы синтеза жира из углеводов (см. рис. 11).
Синтез глицерол-3-фосфата
I этап – при действии соответствующих гликозидаз углеводы подвергаются гидролизу с образованием моносахаридов, которые в цитоплазме клеток включаются в процесс гликолиза. Промежуточными продуктами гликолиза являются фосфодиоксиацетон и 3-фосфоглицериновый альдегид.
IIэтап. Глицерол-3-фосфат образуется в результате восстановления фосфодиоксиацетона – промежуточного продукта гликолиза:
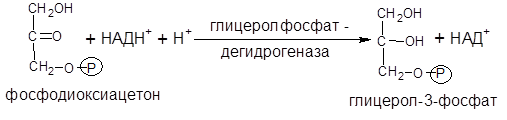
Синтез жирных кислот
Строительным блоком для синтеза жирных кислот в цитозоле клетки служит ацетил-КоА, который образуется двумя путями: либо в результате окислительного декарбоксилирования пирувата. (см. рис. 11, Этап III), либо в результате b-окисления жирных кислот (см. рис. 8).
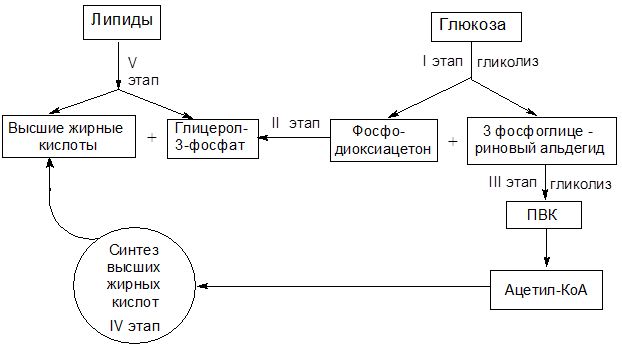
Рисунок 11 – Схема превращения углеводов в липиды
Напомним, что превращения образовавшегося при гликолизе пирувата в ацетил-КоА и его образование при b-окислении жирных кислот происходит в митохондриях. Синтез жирных кислот протекает в цитоплазме. Внутренняя мембрана митохондрий непроницаема для ацетил-КоА. Его поступление в цитоплазму осуществляется по типу облегченной диффузии в виде цитрата или ацетилкарнитина, которые в цитоплазме превращаются в ацетил-КоА, оксалоацетат или карнитин. Однако главный путь переноса ацетил-коА из митохондрии в цитозоль является цитратный (см. рис. 12).
Вначале внутримитохондриальный ацетил-КоА взаимодействует с оксалоацетатом, в результате чего образуется цитрат. Реакция катализируется ферментом цитрат-синтазой. Образовавшийся цитрат переносится через мембрану митохондрий в цитозоль при помощи специальной трикарбоксилаттранспортирующей системы.
В цитозоле цитрат реагирует с HS-КоА и АТФ, вновь распадается на ацетил-КоА и оксалоацетат. Эта реакция катализируется АТФ-цитратлиазой. Уже в цитозоле оксалоацетат при участии цитозольной дикарбоксилат-транспортирующей системы возвращается в митохондриальный матрикс, где окисляется до оксалоацетата, завершая тем самым так называемый челночный цикл:
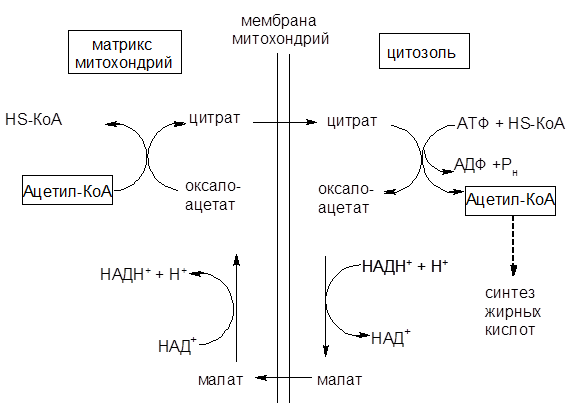
Рисунок 12 – Схема переноса ацетил-КоА из митохондрий в цитозоль
Биосинтез насыщенных жирных кислот происходит в направлении, противоположном их b-окислению, наращивание углеводородных цепей жирных кислот осуществляется за счет последовательного присоединения к их концам двухуглеродного фрагмента (С2) – ацетил-КоА (см. рис. 11, этап IV.).
Первой реакцией биосинтеза жирных кислот является карбоксилирование ацетил-КоА, для чего требуется СО2, АТФ, ионы Mn. Катализирует эту реакцию фермент ацетил-КоА – карбоксилаза. Фермент содержит в качестве простетической группы биотин (витамин Н). Реакция протекает в два этапа: 1 – карбоксилирование биотина с участием АТФ и II – перенос карбоксильной группы на ацетил-КоА, в результате чего образуется малонил-КоА:
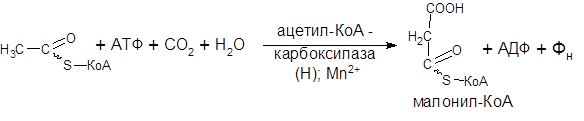
Малонил-КоА представляет собой первый специфический продукт биосинтеза жирных кислот. В присутствии соответствующей ферментной системы малонил-КоА быстро превращается в жирные кислоты.
Нужно отметить, что скорость биосинтеза жирных кислот определяется содержанием сахаров в клетке. Увеличение концентрации глюкозы в жировой ткани человека, животных и повышение скорости гликолиза стимулирует процесс синтеза жирных кислот. Это свидетельствует о том, что жировой и углеводный обмен тесно взаимосвязаны друг с другом. Важную роль здесь играет именно реакция карбоксилирования ацетил-КоА с его превращением в малонил-КоА, катализируемая ацетил-КоА-карбоксилазой. Активность последней зависит от двух факторов: наличия в цитоплазме высокомолекулярных жирных кислот и цитрата.
Накопление жирных кислот оказывает тормозящее влияние на их биосинтез, т.е. подавляют активность карбоксилазы.
Особая роль отводится цитрату, который является активатором ацетил-КоА-карбоксилазы. Цитрат в то же время играет роль связующего звена углеводного и жирового обменов. В цитоплазме цитрат вызывает двойной эффект в стимулировании синтеза жирных кислот: во-первых, как активатор ацетил-КоА-карбоксилазы и, во-вторых, как источник ацетильных групп.
Очень важной особенностью синтеза жирных кислот является то, что все промежуточные продукты синтеза ковалентно связаны с ацилпереносящим белком (HS-АПБ).
HS-АПБ – низкомолекулярный белок, который термостабилен, содержит активную HS-группу и в простетической группе которого содержится пантотеновая кислота (витамин В3). Функция HS-АПБ аналогична функции фермента А (HS-КоА) при b-окислении жирных кислот.
В процессе построения цепи жирных кислот промежуточные продукты образуют эфирные связи с АБП (см. рис. 14):

Цикл удлинения цепи жирных кислот включает четыре реакции: 1) конденсации ацетил-АПБ (С2) с малонил-АПБ (С3); 2) восстановления; 3) дегидротации и 4) второго восстановления жирных кислот. На рис. 13 представлена схема синтеза жирных кислот. Один цикл удлинения цепи жирной кислоты включает четыре последовательных реакции.
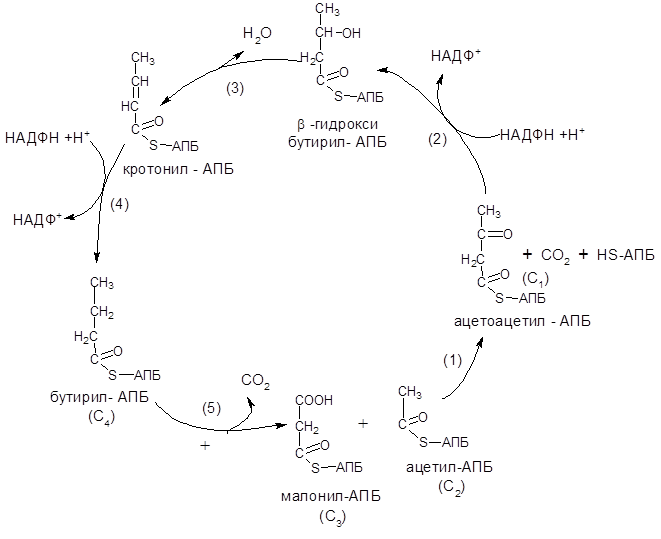
Рисунок 13 – Схема синтеза жирных кислот
В первой реакции (1) – реакции конденсации – ацетильная и малонильные группы взаимодействуют между собой с образованием ацетоацетил-АБП с одновременным выделением СО2 (С1). Эту реакцию катализирует конденсирующий фермент b-кетоацил-АБП-синтетаза. Отщепленный от малонил-АПБ СО2 – это тот же самый СО2, который принимал участие в реакции карбоксилирования ацетил-АПБ. Таким образом, в результате реакции конденсации происходит образование из двух-(С2) и трехуглеродных (С3) компонентов четырехуглеродного соединения (С4).
Во второй реакции (2) – реакции восстановления, катализируемой b-кетоацил-АПБ-редуктазой, ацетоацетил-АПБ превращается в b-гидроксибутирил-АПБ. Восстанавливающим агентом служит НАДФН + Н+.
В третьей реакции (3) цикла-дегидратации – от b-гидроксибутирил-АПБ отщепляется молекула воды с образованием кротонил-АПБ. Реакция катлизируется b-гидроксиацил-АПБ-дегидратазой.
Четвертой (конечный) реакцией (4) цикла является восстановление кротонила-АПБ в бутирил-АПБ. Реакция идет под действием еноил-АПБ-редуктазы. Роль восстановителя здесь выполняет вторая молекула НАДФН + Н+.
Далее цикл реакций повторяется. Допустим, что идет синтез пальмитиновой кислоты (С16). В этом случае образование бутирил-АПБ завершается лишь первый из 7 циклов, в каждом из которых началом является присоединение молекулы молонил-АПБ (С3) – реакция (5) к карбоксильному концу растущей цепи жирной кислоты. При этом отщепляется карбоксильная группа в виде СО2 (С1). Этот процесс можно представить в следующем виде:
С3 + С2 ® С4 + С1 – 1цикл
С4 + С3 ® С6 + С1 – 2 цикл
С6 + С3 ® С8 + С1 –3 цикл
С8 + С3 ® С10 + С1 – 4 цикл
С10+ С3 ® С12 + С1 – 5 цикл
С12 + С3 ® С14 + С1 – 6 цикл
С14 + С3 ® С16 + С1 – 7 цикл
Могут синтезироваться не только высшие насыщенные жирные кислоты, но и ненасыщенные. Мононенасыщенные жирные кислоты образуются из насыщенных в результате окисления (десатурации), катализуруемой ацил-КоА-оксигеназой. В отличие от растительных тканей ткани животных обладают весьма ограниченной способностью превращать насыщенные жирные кислоты в ненасыщенные. Установлено, что две наиболее распространенные мононенасыщенные жирные кислоты – пальмитоолеиновая и олеиновая – синтезируются из пальмитиновой и стеариновой кислот. В организме млекопитающих, в том числе и человека, не могут образовываться, например, из стеариновой кислоты (С18:0) линолевая (С18:2) и линоленовая (С18:3) кислоты. Эти кислоты относятся к категории незаменимых жирных кислот. К незаменимым жирным кислотам относят также арахиновую кислоту (С20:4).
Наряду с десатурацией жирных кислот (образование двойных связей) происходит и их удлинение (элонгации). Причем, оба эти процесса могут сочетаться и повторяться. Удлинение цепи жирной кислоты происходит путем последовательного присоединения к соответствующему ацил-КоА двууглеродных фрагментов при участии малонил-КоА и НАДФН+Н+.
На рисунке 14 представлены пути превращения пальмитиновой кислоты в реакциях десатурации и элонгации.
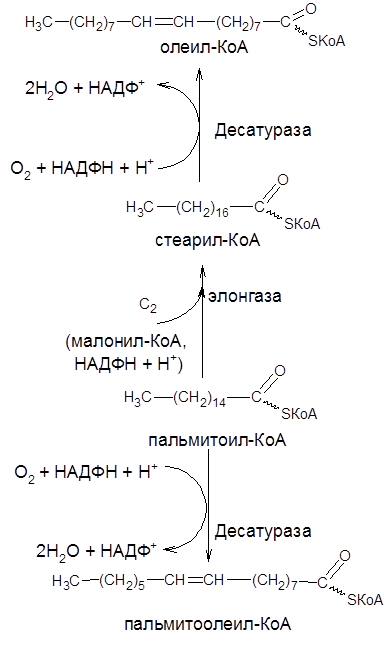
Рисунок 14 – Схема превращения насыщенных жирных кислот
в ненасыщенные
Завершается синтез любой жирной кислоты отщеплением HS-АПБ от ацил-АПБ под влиянием фермента деацилазы. Например:
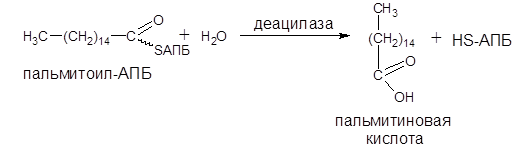
Далее жирная кислота подвергается активации.