Окисление ПВК до ацетил-КоА происходит при участии ряда ферментов и коферментов, объединенных структурно в мультиферментную систему, получившую название «пируватдегидрогеназный комплекс».
Процесс окислительного декарбоксилирования ПВК происходит в матриксе митохондрий. В нем принимают участие (в составе пируватдегидрогеназного комплекса) 3 фермента (пируватдегидрогеназа, дигидролипоилдегидрогеназа и дигидролипоилацетилтрансфераза) и 5 коферментов (тиаминпирофосфат (ТПФ (витамин В1)), амид липоевой кислоты, коэнзим А (СоА (витамин В3)), ФАД (флавинадениндинуклеотид (витамин В2)),НАД+(никотинамидадениндинуклеотид (витамин РР)).
Суммарную реакцию, катализируемую пируватдегидрогеназным комплексом, можно представить следующим образом:

Образовавшийся в процессе окислительного декарбоксилирования ацетил-СоА окисляется далее в цикле трикарбоновых кислот с образованием СО2 и АТФ.
27. Цикл трикарбоновых кислот (Цикл Кребса).
Этот цикл известен также под названием цикла лимонной кислоты или цикла Кребса в честь Г. Кребса, определившего последовательность реакций цикла. За это открытие Кребс получил в 1953 году Нобелевскую премию.
Хотя цикл трикарбоновых кислот мы рассматриваем во взаимосвязи с катаболизмом углеводов, однако его роль в метаболизме веществ гораздо шире.
Цикл Кребса выполняет следующие функции:
1. Интегративную – объединяет пути катаболизма углеводов, жиров, белков; во всех аэробных организмах он выступает в роли центрального метаболического пути углерода.
2. Амфиболическую – выполняет не только катаболическую функцию распада ацетильных остатков, но и анаболическую, поскольку субстраты цикла используются для синтеза других веществ;
3. Энергетическую – совместно с цепью переноса электронов является основным поставщиком химической энергии в форме АТФ.
Реакции цикла протекают в матриксе митохондрий и представляют восемь последовательных превращений.
Первая реакция цикла начинается с конденсации ацетил-СоА с оксалоацетатом, в результате чего образуется лимонная кислота.
Реакция катализируется цитрат-синтазой (класс лиазы):

В результате второй реакции образовавшаяся лимонная кислота подвергается дегидрагированию с образованием цис-аконитовой кислоты, которая, присоединяя молекулу воды, переходит в изолимонную (изоцитрат). Обратимые реакции гидратации-дегидратации катализирует фермент аконитаза (класс лиазы):

Третья реакция лимитирует скорость цикла Кребса. Изолимонная кислота дегидрируется в присутствии НАД–зависимой дегидрогеназы (класс оксидоредуктазы):

Во время четвертой реакции происходит окислительное декарбоксилирование a-кетоглутаровой кислоты с образованием высокоэнергетического соединения сукцинил-СоА.
Механизм этой реакции сходен с механизмом реакции окислительного декарбоксилирования ПВК. a-кетоглутаратдегидроге-назный комплекс напоминает по своей структуре пируватдегидрогеназный комплекс (см. пируватдегидрогеназный комплекс):

Пятая реакция катализируется ферментом сукцинил-СоА-синтетазой (класс лигазы):

Данная реакция является реакцией субстратного фосфорилирования, т.к. за счет высокоэргической тиоэфирной связи сукцинил-КоА происходит образование высокоэргической фосфатной связи ГТФ.
В результате шестой реакции сукцинат дегидрируется в фумаровую кислоту. Окисление сукцината катализируется сукцинатдегидрогеназой (класс оксидоредуктазы), простетической группой при этом является ФАД.

Седьмая реакция осуществляется под влиянием фермента фумаратгидратазы (класс лиазы):

В ходе восьмой реакции цикла трикарбоновых кислот под влиянием НАД-зависимой малатдегидрогеназы происходит окисление яблочная кислоты в оксалоацетат:

Таким образом за один оборот цикла происходит полное окисление одной молекулы ацетил-СоА и регенерация оксалоацетата, который вновь может конденсироваться с новой молекулой ацетил-СоА.
28. Дыхательная цепь переноса электронов. Окислительное фосфорилирование, биосинтез АТФ
Для непрерывной работы цикла Кребса необходимо постоянное поступление в систему ацетил-СоА, а восстановленные коферменты НАДН и ФАДН2 должны снова и снова окисляться. НАДН и ФАДН2 окисляются молекулярным кислородом не сразу и непрямо (в реакциях цикла Кребса молекулярный кислород нигде не участвует, хотя этот цикл является аэробным, т.е. кислородзависимым). Процесс идет постепенно с помощью последовательно функционирующих окислительно-восстановительных систем, переносящих водород, отнятый у субстрата в виде эквивалента электронов и протонов к молекулярному кислороду. Совокупность этих систем получила название дыхательной цепи переноса электронов.
Ферментные системы, составляющие дыхательную цепь переноса электронов, встроены во внутренней мембране митохондрий.

Митохондрии – это органеллы клетки (рис.3), играющие роль «силовых станций» клетки. Данная органелла является местом, где протекает цикл трикарбоновых кислот и окисление жирных кислот, генерирующие восстановленные коферменты, которые затем окисляются с помощью дыхательной электронтранспортной цепи.
Митохондрия окружена двумя мембранами с сильно различающимися свойствами. Внутренняя мембрана митохондрии является барьером для протонов. Она способна пропускать такие молекулы как вода, молекулярный кислород, СО2, но фактически непроницаема для большинства полярных и ионизированных веществ (Na+, K+, НАД+, НАДН и т.д.). Наружная мембрана свободно проницаема для соединений с молекулярной массой около 10.000.
29. Компоненты дыхательной цепи распределены по плоскости внутренней мембраны равномерно. Они находятся в липидном окружении, прочно связаны с мембраной.
Также в митохондриальной мембране находится АТФ-синтетазный комплекс, ответственный за образованием АТФ.
Синтез АТФ из АДФ и фосфорной кислоты, который происходит с использованием энергии, освобождающейся при окислении веществ в живых клетках, и сопряжен с переносом электронов по дыхательной цепи, называется окислительным фосфорилированием.
Дыхательная цепь переноса электронов включает пять комплексов белков, в состав которых входят:
1. пиридинзависимые дегидрогеназы, коферментом которых является НАД+

2. флавинзависимые дегидрогеназы, коферментом которых является ФАД или ФМН

3. небелковый переносчик убихинон, который называют коферментом
Q (KoQ).

4. Цитохромная система.
Цитохромы представляют собой гемопротеины, т.е. белки, содержащие в качестве простетической группы – гем. В составе гема имеется ион железа, который при переносе электронов подвергается обратимому окислению-восстановлению.

Цитохромы относятся к одноэлектронным окислительно-восстановительным соединениям. Они не могут принимать или отдавать водород, поэтому их называют переносчиками электронов.
Цитохромы подразделяются на цитохромы а, в, с и d в соответствии с положением их спектральных полос поглощения в восстановленном состоянии. Гемовые простетические группы цитохромов также имеют небольшие структурные различия; кроме того они отличаются структурой белка, с которым связан гем. Цитохромы а и а3 существуют и функционируют в виде единого прочного комплекса. Особенностью комплекса а×а3 является наличие в нем меди, связанной с компонентом а3, необходимой для его функционирования.
Перенос электронов комплексом а×а3 включает следующие реакции:

Особенностью комплекса а×а3 является то, что эта система цитохромов непосредственно реагирует с молекулярным кислородом воздуха, т.е. электроны передаются на кислород, переводя его в ионы. Комплекс а×а3 называют цитохромоксидазой.
5.Железосерные белки, или негеминовые белки, т.е. белки, в составе которых железо не связано с гемом.
Компоненты дыхательной цепи расположены по мере возрастания окислительно-восстановительного потенциала. Окислительно-восстанови-тельной потенциал пары НАД/НАДН + Н+ = -0,32 В, а пары О2/Н2О = +0,82 В.
Последовательность реакций, протекающих в дыхательной цепи переноса электронов представлена в виде схемы (рис. 4).

Процесс начинается с переноса электронов и протонов от восстановленного субстрата АН2 к НАД+ (например: малат + НАД+ ® оксалоацетат + НАДН + Н+ (см. реакции цикла Кребса)).
Восстановленный НАДН + Н+ передает два электрона и два протона ФМН. От флавина электроны и протоны переходят к убихинону (CoQ). После убихинона пути электронов и протонов расходятся. Электроны в определенной последовательности передаются по цепи цитохромов и, наконец, через цитохромоксидазу (а×а3) достигают кислорода воздуха, переводя его в ион. При этом осуществляется одноэлектронный перенос по системе цитохромов. Существует также укороченная дыхательная цепь, когда субстрат А1Н2 взаимодействует с ФАД-дегидрогеназой (например: янтарная кислота + ФАД ® фумаровая кислота + ФАДН2 (см. реакции цикла Кребса)), которая восстанавливается до ФАДН2 и далее электроны и протоны идут на CoQ. Дальнейший путь электронов и протонов совпадают.
Протоны после убихинона выталкиваются в межмембранное пространство митохондрии за счет энергии, извлекаемой при движении электронов по дыхательной цепи.
В результате наружная сторона внутренней мембраны заряжается положительно, а матрикс митохондрии – отрицательно. Возникает электрохимический градиент концентраций ионов ΔМН+.

Диффузия протонов обратно через мембрану, т.е. в область с их меньшей концентрацией, происходит по концентрационному градиенту и сопряжена с фосфорилированием, осуществляемым АТФ-синтетазой.
Согласно хемиосмотической теории, разработанной П. Митчелом, энергия, сосредоточенная в протонном потенциале, используется АТФ-синтетазой для выполнения химической работы, т.е. синтеза АТФ:
АДФ + Н3РО4 ® АТФ + НОН
В данном случае синтез АТФ происходит одновременно за счет энергии, выделяемой по мере продвижения электронов по дыхательной цепи, т.е. одновременно с окислением водорода, снятого с субстрата до воды, синтезируется АТФ. Такой способ синтеза АТФ называется окислительным фосфорилированием.
В дыхательной цепи в трех пунктах происходит сопряжение переноса электронов (окисления) и фосфорилирования. Первым этапом является перенос электронов от восстановленной анаэробной дегирогеназы НАДН + Н+ к флавиновой дегидрогеназе ФМН. Второй точкой сопряжения является переход электронов от цитохрома в на цитохром с1. Третья молекула АТФ выделяется, когда цитохромоксидаза (а×а3) передает электроны кислороду воздуха. Таким образом, при окислении молекулы водорода, снятой с субстрата НАД-зависимой дегидрогеназой, синтезируется 3 молекулы АТФ. Если водород передается ФАД-зависимой дегидрогеназой, то первая точка сопряжения минуется, поскольку от ФАДН2 электроны и протоны передаются коферменту Q, следовательно образуется только 2 молекулы АТФ.
30. Пентозофосфатный путь окисления углеводов
Помимо катаболизма углеводов по пути гликолиза существует другой, альтернативный путь, получивший название гексозомонофосфатного или пентозофосфатного пути.
Расхождение путей окисления начинается со стадии образования глюкозо-6-фосфата.
Если глюкозо-6-фосфат изомеризуется во фруктозо-6-фосфат, который фосфорилируется второй раз и превращается во фруктозо-1,6-дифосфат, то в этом случае дальнейший распада углеводов происходит по обычному гликолитическому пути. Если второго фосфорилирования фруктозы-6-фосфат не происходит, то фосфорилированная глюкоза может подвергаться прямому –окислению до фосфопентоз.
В норме доля пентозофосфатного пути в количественном превращении глюкозы обычно невелика, варьирует у разных организмов и зависит от типа ткани, ее функционального состояния.
Значение этого пути в обмене веществ велико. Этот путь выполняет 3 основные функции:
1) поставляет НАДФН, который используется в биосинтезе жирных кислот, холестерина и т.д.;
2) является источником рибозо-5-фосфата, необходимого для синтеза нуклеиновых кислот;
3) генерирует эритрозо-4-фосфат, который используется для синтеза ароматических аминокислот: фенилаланина, тирозина, триптофана.
Реакции пентозофосфатного цикла протекают в цитоплазме клетки и состоят из окислительной и неокислительной стадий.
Первая окислительная реакция – дегидрирование глюкозо-6-фосфата:
 6-Фосфоглюконолактон – соединение неустойчивое и с большой скоростью гидролизуется:
6-Фосфоглюконолактон – соединение неустойчивое и с большой скоростью гидролизуется:

Во второй окислительной реакции 6-фосфоглюконовая дегидрируется и декарбоксилируется:
 Рибулозо-5-фосфат может изомеризоваться с образованием ксилулозо-5-фосфат или рибозо-5-фосфат:
Рибулозо-5-фосфат может изомеризоваться с образованием ксилулозо-5-фосфат или рибозо-5-фосфат:
 При определенных условиях пентозофосфатный путь на этом этапе может быть завершен. Однако при других условиях наступает неокислительный этап пентозофосфатного цикла. Реакции этого этапа не связаны с использованием кислорода и протекают в анаэробных условиях.
При определенных условиях пентозофосфатный путь на этом этапе может быть завершен. Однако при других условиях наступает неокислительный этап пентозофосфатного цикла. Реакции этого этапа не связаны с использованием кислорода и протекают в анаэробных условиях.
Шесть молекул глюкозо-6-фосфата, вступая в пентозофосфатный цикл, образуют 6 молекул рибулозо-5-фосфата и 6 молекул СО2, после чего из 6 молекул рибулозо-5-фосфата снова регенерируется 5 молекул глюкозо-6-фосфата. Образовавшийся НАДФН используется в цитозоле на восстановительные синтезы и, как правило, не участвует в окислительном фосфорилировании, протекающем в митохондриях.

Фотосинтез
В растительных клетках глюкоза синтезируется за счет фотосинтеза.
Фотосинтез – это синтез углеводов и других органических соединений из неорганических – СО2 и Н2О – под действием энергии солнечного света.
Различают две части фотосинтеза – световой и темновой процессы.
Световой процесс включает поглощение света, разложение воды на водород и кислород (фотоокисление),восстановление НАДФ+, фосфорилирование АДФ:


Темновой процесс фотосинтеза – это восстановление диоксида углерода за счет водорода НАДФН и энергии АТФ. 
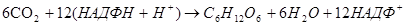

Суммарный результат светового и темнового процессов:

Выделяемый растениями кислород образуется из воды при ее разложении в световом процессе.