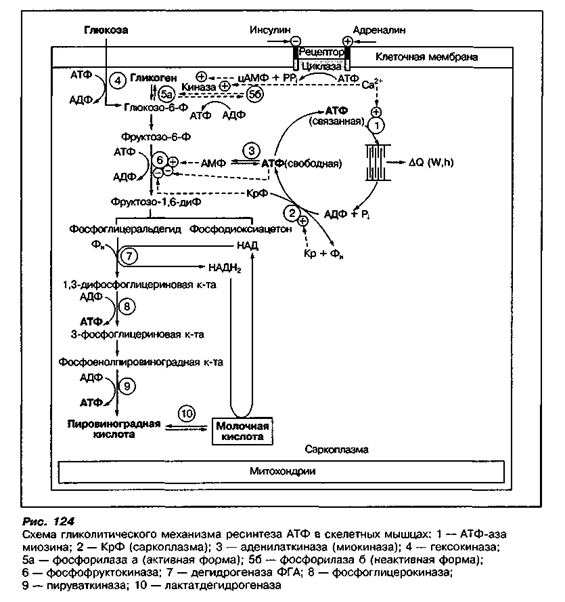
Как только в процессе анаэробной мышечной работы креатинфосфокиназный механизм перестает обеспечивать необходимую скорость восстановления АТФ в мышцах, в энергообеспечение работы вовлекается анаэробный гликолитический механизм ресинтеза АТФ. В процессе гликолиза используются в основном внутримышечные запасы гликогена, а также глюкоза, поступающая из крови. Они постепенно расщепляются до молочной кислоты с участием многих ферментов (рис. 124).
Большинство ферментов гликолиза локализовано в саркоплазме мышечных волокон. Ферменты фосфорилаза и гексокиназа, обеспечивающие начальные реакции гликолиза, легко активируются при повышении концентрации АДФ и неорганического фосфата в саркоплазме. Кроме того, образование активной формы фосфорилазы стимулируется катехоламинами и ионами Са2+, уровень которых повышается при мышечном сокращении. Все это способствует быстрому подключению гликолиза к ресинтезу АТФ уже с первых секунд работы, о чем свидетельствует повышение концентрации молочной кислоты в мышцах.
|
Активации гидролиза способствует также снижение концентрации креатинфосфата в мышцах и накопление АМФ, образующегося в миокиназной реакции ресинтеза АТФ.
Обобщенно процесс гликолиза и гликогенолиза может быть представлен в виде следующих уравнений:
С6Н1208+ 2АДФ + 2Н3Р04 → 2С3Н6О3 + 2АТФ + 2Н2О
Глюкоза Молочная кислота
[С6Н1005]n + 3АДФ + 3Н3Р04 → 2С3Н603 + [С6Н1005]n-1 + 3АТФ + 2Н2О
Гликоген Молочная кислота
Энергетический баланс гликолиза в случае, когда исходным веществом служит глюкоза, составляет 2 моля АТФ на 1 моль расщепляемых углеводов, а гликогенолиза, когда исходным веществом является гликоген мышц, — 3 моля АТФ на 1 моль расщепляемого глюкозного эквивалента.
Максимальная мощность гликолиза у хорошо тренированных спортсменов может составлять 3,1 кДж • кг -1 • мин -1, а у нетренированных людей — 2,5 кДж • кг -1• мин -1. Это несколько ниже, чем мощность креатинфосфокиназной реакции, но в 2—3 раза выше мощности аэробного процесса.
На максимальную мощность этот механизм выходит уже на 20—30-й секунде после начала работы, т. е. его скорость развертывания значительно меньше, чем креатинфосфокиназного. К концу 1 -и минуты работы гликолиз становится основным механизмом ресинтеза АТФ. Однако при дальнейшей работе наблюдается снижение активности ключевых ферментов гликолиза под влиянием образующейся молочной кислоты или снижения внутриклеточного рН, что приводит к снижению скорости ресинтеза АТФ в этом механизме.
Метаболическая емкость гликолиза, определяемая внутримышечными запасами углеводов и зависящая от резервов буферных систем, стабилизирующих величину внутриклеточного рН, обеспечивает поддержание анаэробной работы продолжительностью от 30 с до 2—6 мин.
Общее количество энергии, которое образуется в гликолитическом механизме у нетренированных людей, не превышает 840 Дж • кг -1, что соответствует концентрации молочной кислоты в крови около 13 ммоль • л -1, которая для них является граничной. У спортсменов, которые в процессе тренировки используют физические нагрузки анаэробной гликолитической направленности, лактатная емкость составляет 1760—2090 Дж • кг -1, что соответствует наличию молочной кислоты в крови в пределах 25— 30 ммоль • л -1. Тем не менее у спортсменов, специализирующихся в видах спорта на выносливость, после выполненной мышечной работы концентрация молочной кислоты в крови не превышает 10—13 ммоль • л -1 и зависит от содержания быстросокращающихся волокон в скелетных мышцах.
Гликолитический механизм ресинтеза АТФ отличается невысокой эффективностью, так как в процессе анаэробного распада 1 моля глюкозы из 2880 кДж энергии, содержащейся в ней, высвобождается только 240 кДж. Большая часть энергии остается в молекулах образующейся молочной кислоты и может быть выделена только путем аэробного окисления.
Из общего количества энергии, выделенной в ходе гликолиза, в доступную для использования форму макроэргических фосфатных связей АТФ преобразуется от 80 до 125 кДж, а остальная энергия выделяется в виде тепла. Исходя из этого метаболическая эффективность гликолиза оценивается значениями КПД порядка 0,35—0,52. Это означает, что примерно половина всей выделяемой энергии превращается в тепло и не может быть использована в работе. В результате при гликолитической работе повышается скорость теплопродукции в работающих мышцах и их температура до 41-42•С.
Гликолиз - основной путь энергообразования в упражнениях субмаксимальной мощности, предельная продолжительность которых составляет от 30 с до 2,5 мин (бег на средние дистанции, плавание на 100 и 200 м, велосипедные гонки на треке и т. п.).
За счет гликолиза совершаются длительные ускорения по ходу упражнения и на финише дистанции.
Гликолитический механизм энергообразования является биохимической основой специальной скоростной выносливости организма.
Гликолитический путь ресинтеза АТФ сопровождается накоплением молочной кислоты в мышцах, концентрация которой находится в прямой зависимости от мощности и общей продолжительности упражнения.
Молочная кислота в водной среде подвергается диссоциации на ионы и вызывает изменение концентрации водородных ионов и величины рН внутриклеточной среды:
Н2О
СНз-СНОН-СООН ← → СНз-СНОН-СОо - + Н+
Молочная кислота Ион молочной кислоты (лактат)
Умеренный сдвиг рН в кислую сторону активирует работу ферментов дыхательного цикла в митохондриях и усиливает аэробное энергообразование. Значительное изменение рН среды в мышцах от 7,1 в состоянии покоя до 6,5 при изнеможении угнетает ферменты, регулирующие скорость гликолиза и сокращение мышц. При значении внутримышечного рН 6,4 прекращается расщепление гликогена, что вызывает резкое снижение уровня АТФ и развитие утомления.
Увеличение количества молочной кислоты в саркоплазме мышц сопровождается изменением осмотического давления. При этом вода из межклеточной среды поступает внутрь мышечных волокон, вызывая их набухание, что может привести к сдавливанию нервных окончаний и возникновению болевых ощущений в мышцах.
Молочная кислота может диффундировать через клеточные мембраны по градиенту концентрации и поступать из работающих мышц в кровь. Обычно максимальное накопление молочной кислоты в крови наблюдается через 5—7 мин после работы. Молочная кислота взаимодействует с бикарбонатной буферной системой крови, что приводит к образованию "неметаболического" избытка СО2:
СНз-СНОН-СООН + NаНСОз → СН3-СНОН-СООNа + Н+ + НСО3 -
Н+ + НСО3 - → Н2СО3 → Н2О + СО2 ↑
Увеличение концентрации водородных ионов и повышение напряжения С02 в крови способствуют активации дыхательного центра, поэтому при выходе молочной кислоты в кровь резко усиливается легочная вентиляция и поставка кислорода к работающим мышцам. Значительное накопление молочной кислоты, появление избыточного СО2, изменение рН и гипервентиляция легких, отражающие усиление гликолиза в мышцах, обнаруживается при увеличении интенсивности выполняемого упражнения более 50 % максимальной аэробной мощности. Этот уровень нагрузки обозначается как порог анаэробного обмена (ПАНО), или порог лактата (ПЛ). Чем раньше он будет достигнут, тем быстрее вступит в действие гликолиз, сопровождающийся накоплением молочной кислоты и последующим развитием утомления работающих мышц.
Величина ПАНО является важным показателем эффективности процессов энергообразования в мышцах, интенсивности тренирующих нагрузок, роста степени тренированности, который широко используется при биохимическом контроле функционального состояния спортсмена. Наиболее обоснованным является метод построения индивидуальных кривых зависимости концентрации лактата от мощности выполняемой физической работы, которую выражают либо в ваттах (Вт), либо в процентах от МПК. Резкий излом кривой свидетельствует о переходе мышц в анаэробный режим работы, после чего концентрация молочной кислоты с увеличением мощности нагрузки будет увеличиваться за счет повышения интенсивности гликолиза.
У многих спортсменов ПАНО соответствует уровню молочной кислоты, равному 2—4 ммоль • л-1. С ростом степени тренированности на выносливость порог лактата увеличивается, т. е. наступает при более интенсивной работе.