
Развитие кист ткани в пробирке было зарегистрировано более 40 лет назад (Hogan и др, 1960;. Matsukayashi и Akao, 1963); Однако морфологическое сходство bradyzoites и тахи- zoites с помощью световой микроскопии затрудняет изучение этих дифференциации событий, пока выработка антител не брадизоитный-специфические антигены.
В 1977 году, Hoff показал, что в пробирке производимая кист привело к ооцитов экскреции у кошек с prepatent периода, который согласуется с кист ткани (т.е. созревают bradyzoites) (Hoff и др., 1977). С помощью ТЭМ, было показано, что в то время как киста-подобные структуры присутствовали в течение 3 дней инфицирования клеток-хозяев в культуре ткани, с помощью биоанализа (у кошек) зрелые цисты не присутствовали до 6 дней после инфицирования клеток-хозяев в пробирке (Dubey, 1997). Продолжительное прохождение Т. гондий или другого Apicomplexa в пробирке, может привести к потере их способности отличаются-entiate в другие стадии. Так, например, длительное прохождение Besnoitia jellisoni в пробирке приводит к потере его способности образовывать цисты ткани у мышей, и многие изолят II типа (например, PLK) Т. гондий не может образовывать ооцист у кошек (Френкель и др., 1976),
Брадизоитные-специфический моноклональные антитела (таблица 13.1) значительно способствовали исследованиям брадизоитного развития в пробирке и Понимать-ции методов индукции дифференциального-ции. В последнее время, паразит линия также была разработана, которые используют GFP под контролем промоторов Vari-OUs стадии специфической (Синг и др., 2002; Мо и др., 2004). Используя паразиты, содержащие Chlo-ramphenicol ацетилтрансферазы (CAT) выражается конститутивно отα-tubulin промотор (TUB1), и βгалактозидазы (βGAL) экспрессируется из брадизоитного-специфического промотора (BAG1), можно измерить как скорость роста и степень дифференцировки брадизоитной (Итон и др., 2005).
В исследованиях ткани культуры, то очевидно, что Брейди zoites спонтанно конвертировать в тахизоитами и тахизоиты спонтанно конвертировать в Брейди zoites (Jones и др, 1986;.. Линдсей и др, 1991, 1993а, 1993b;. Bohne и др 1993, 1994; Макью и др, 1993;. Soete и др, 1993, 1994;. Попель и соавт, 1994, 1996;. Вейс и соавт, 1994;. Parmley и др., 1995; Lane и др., 1996; Soete и Dubremetz, 1996;. De Champs и др, 1997). Было отмечено Matsukayashi в 1963 году, и была подтверждена несколькими группами с использованием брадизоитный-специфических моноклональных антител, что Т. гондий штаммы с более медленной скоростью репликации были более склонны к развитию кист в пробирке, и что замедляет репликацию вирулентных штаммов позволит ткани цисты вирулентных пятен развивать в пробирке (Matsukayashi и Akao, 1963). штаммы с низкой вирулентности являются высокими кистой образующих штаммов в

| РАЗВИТИЕ ТКАНЕЙ кист и BRADYZOITES IN VITRO | |||||
| Таблица 13.1 Маркеры Общие брадизоитный | |||||
| Размер по | |||||
| Имя | моноклональные антитела | иммуноблот | Расположение по | Предложил | |
| антиген | антитело | (КД) | ЕСЛИ | функция | клонированный |
| BAG1 / hsp30 | 7E5 | цитоплазма | Малый тепло- | да | |
| ака BAG5 | 74.1.8 | белок шока | |||
| BSR4 (p36) | T84A12 | поверхность | семья SAG1 | да | |
| антигена, а также в | |||||
| спорозоиты | |||||
| SAG4A (P18) | T83B1 | поверхность | поверхностный антиген | да | |
| Никто | DC11 | Не | поверхность | ? | Один из SAG |
| реактивный | семья? | ||||
| p21 | T84G11 | поверхность | поверхностный антиген | Один из SAG | |
| семья? | |||||
| p34 | T82C2 | поверхность | ? | Один из SAG | |
| семья? | |||||
| SRS9 | мышиный | поверхность | Поверхностный антиген, | да | |
| поликлональные | SAG, связанные с | ||||
| только | последовательность | ||||
| MAG1 | Никто | матрица | Да, переработанное | ||
| исследования | |||||
| указывают также | |||||
| выражено в | |||||
| тахизоиты | |||||
| Никто | E7B2 | матрица | нет | ||
| Никто | 93,2 | Не | матрица | нет | |
| реактивный | |||||
| Никто | 1.23.29 | матрица | нет | ||
| CST1 | 73,18; также | Киста стены | нет | ||
| признаны | |||||
| DBA лектин | |||||
| CST1? | СС2 | Киста стены | То же самое CST1? | нет | |
| (Брадизоитный) | |||||
| 40 (тахизоит) | |||||
| LDH2 | поликлональные сыворотки | 35 кДа | цитозольного | гликолиз | Да, |
| слабо кросс- | (33 кДа для | тахизоит | |||
| реагирует на LDH1 | LDH1) | изоформа | |||
| LDH1 | |||||
| ENO1 | поликлональные сыворотки | Ядерная и | гликолиз | Да, | |
| в ENO2 и | цитозольная | тахизоит | |||
| ENO1 не | изоформы | ||||
| перекрестно реагируют | ENO2 | ||||
346 РАЗВИТИЕ брадизоитного
мышей (например, тип II, такие, как ME49 или Pru, или типа III штаммов 76K или Вег), и имеют более высокую Спонта-ое скорость образования кист в культуре, чем делают вирулентный тип штаммы I, такие как RH (Soete и соавт., 1994). Этот биологический фенотип также коррелирует с узорами экспрессии генов, наблюдаемой (Radke и др., 2005). Кроме того, мы обнаружили, что эффективность производства цист на авирулентном Т. гондий изолирует уменьшается с продлены в пробирке прохода (это коррелирует с развитием более быстрого ростом фенотипа этих штаммов в длительной культуре ткани).
стрессовые условия связаны с induc Тион развития брадизоитного - то есть больше bradyzoites в этих условиях, чем можно было бы ожидать от простого ингибирования репликации тахизоит. Условия, которые индуцируют брадизоитный образование внутри клеток-хозяев вспыльчивость-ature стресс (43°C;. Soete и др, 1994), рН стресса (рН 6,6-6,8 или 8,0-8,2; Soete и др, 1994;. Вейс и др., 1995; Soete и Dubremetz, 1996), химический стресс (Na Арсенит; Soete др., 1994), и питательные напряжения (аргинин голодание; Фокс и др, 2004).. В мышиных линиях макрофагов, полученных из костного мозга, IFN- &γповышает экспрессию антигена брадизоитной (Bohne и др., 1994). Это по-видимому, из-за производство оксида азота (NO), так как брадизоитная различаются-entiation ингибировались обработка макрофагов с NMMA (Nг-monomethyl-L-аргинин, ингибитор синтазы индуцируемый азотно-оксид) (Bohne и др., 1994). Брадизоитный дифференциация также повышается за счет нитропруссид натрия (SNP), экзогенного донора NO (Bohne и соавт, 1994; Weiss. И др, 1996;. Киркман и др., 2001). Точно так же, как олигомицин (ингибитор митохондриальной АТФ-синтетазы функции) и антимицин А (ингибитор переноса электронов дыхательной цепи) увеличивает экспрессию антигена брадизоитной (Bohne и соавт, 1994;. Tomavo и Boothroyd, 1995). рН стресс является одним из наиболее часто используемых методов индуцируют дифференцировку брадизоитной в лаборатории (рис 13.2).
Вклад клетки-хозяина на стадии CONVER-сии еще предстоит выяснить. Две группы сообщили (Вайс и др., 1998; Yahiaoui и др., 1999), что воздействие внеклеточного тахизоитов к стрессовым условиям (рН 8,1) приведет к увеличению
брадизоитный дифференциация, в соответствии с прямым воздействием стресса на паразита. Тем не менее, большинство агентов, которые индуцируют дифференцировку оказывают глубокое воздействие на клетки-хозяева, и вполне вероятно, что изменения в передаче сигналов клетки-хозяина также имеют существенное влияние на брадизоитный дифференциации. Недавно было показано, что паразит-специфический ингибитор PKG соединение 1 (Дональд и Освободитель, 2002; Дональда и др., 2002; Gurnett и др., 2002;. Нарэ и др, 2002) требует синтеза хост-клеточного белка для того, чтобы вызвать образование брадизоитный, предполагая, что его действие на дифференцировку опосредовано через возмущение клетки-хозяина, а не непосредственно на паразита (Radke и др., 2006).
Когда клетки заражены bradyzoites из цистой ткани, дифференциаций в тахизоиты и появление тахизоят антигены (SAG1) происходит в течение 15 часов и до клеточного деления произошло (Soete и Dubremetz, 1996). Вакуоли содержащих организмы, экспрессирующие только тахизоят антигены отчетливо видны в течение 48 часов после заражения. Когда брадизоитный дифференциация происходит в культуре клеток после инфицирования тахизоитов, все доступных в настоящее время маркеров для формирования брадизоитного, за исключением p21 (мАт T84G10), может быть обнаружено в течение 24 часов инфекции (Gross и др., 1996;. Лейн и соавт, 1996). Это включает в себя маркеры брадизоитный поверхностных антигенов, а также те, которые связаны с образованием кисты стенки. Преобразование между этими двумя этапами является быстрым событием,
К 3-х дней после воздействия условий, которые вызывают развитие брадизоитный, вакуоли присутствуют в культуре ткани с электронными микро-скопическим характеристик кист; Однако реакционная способность к МАБ T84G10 (p21) не появляется до тех пор, в течение 5 дней. По оценке биоанализом у кошек, зрелые / функ-ционных цисты не не образуются до тех пор, по крайней мере, 6 дней в культуре. Дополнительные маркеры зрелых функциональных кист необходимы, чтобы облегчить в лабораторных исследованиях по кисте биологии. Там может быть различие в экспрессии антигенов и репликации в начале bradyzoites и конце bradyzoites, но детальная хронология экспрессии маркеров до сих пор не разработана.
| Клеточный цикл И РАЗВИТИЕ брадизоитного |
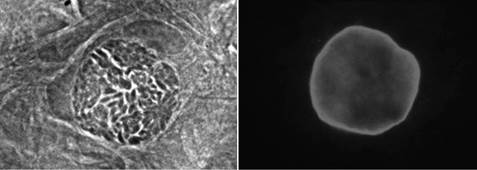
A В

С D
Рисунок 13.2 Развитие Т. Gondii этапы жизни в пробирке.
Первичные мышиные астроциты (рис 13.2a, 13.2B) и фибробласты крайней плоти человека (фиг 13.2C, 13.2d), инфицированных T. гондий (ME49 / PLK штамма). Человеческие фибробласты, зараженные Т. гондий (ME49 / PLK штамма) поддерживали при рН 8,0 (рис 13.2C) или рН 7,2 (рис 13.2d) в течение 3 дней.
(A) Фаза микроснимок демонстрируя в пробирке кисту ткани.
(B) ИФА с использованием анти-CST1 (MAB 73.18), демонстрирующее окрашивание кисты стенки брадизоитный пара-sitophorous вакуоли (соответствующая фазовая микроскопия показана на рисунке 13.2a).
(C) ИФ с использованием анти-BAG1 (монАТа 74.1.8) демонстрирует брадизоитное развитие в нескольких паразитофорной вакуоле. Окрашивание происходит по всей цитоплазме bradyzoites.
(D) ИФ с использованием анти-SAG1 (MAB DG52), демонстрирующими тахизоят развитие и формирование розетки. Окрашивание локализуется на поверхности каждого тахизоит.
Эта цифра воспроизводится в цвете в разделе Цвет пластины.
Клеточного цикла и
РАЗВИТИЕ брадизоитного
Вполне вероятно, что брадизоитное отличие от тахизоитов является программируемым ответом, связанный с замедлением репликации и удлинения клеточного цикла (Иероним и др., 1998), аналогично запрограммированным расширением и дифференциации сообщалась в других
кокцидий. Клеточный цикл в тахизоитах является символом-теризуется основными G1 и S фазами и относительно коротким G2+М. Как замедляет Т. гондий репликации, происходит увеличение продолжительности фазы G1 клеточного цикла. Не известно ли контрольные точки в пределах этого клеточного цикла отличаются от наблюдаемых в дрожжах и в клетках млекопитающих, но предварительные шпилька-х годов предложить некоторые различия (Radke и соавт., 2001;
348 РАЗВИТИЕ брадизоитного
Хан и др., 2002). Когда Т. гондий спорозоиты (VEG) штамм инфицируют фибробластов человека в пробирке, они превращаются в быстро делящихся тахизоиты, имеющий период полураспада 6 часов. После 20 делений (Approxi-зительно 5 дней в культуре), эти тахизоиты переход к более медленной скорости роста с периодом полураспада 15 часов (Radke и др., 2001). Брадизоитные дифференциации, как определена выражением брадизоитных специфических антигенов, происходят спонтанно, когда-Тион популярного переходит к более медленной скорости роста (т1/2 16 ч), но не виден в быстро разделяющей (т1/26 ч) организмы. Анализ экспрессии генов с SAGE показывает, что уровни мРНКа брадизоитных хорошо коррелируют с тем, что было обнаружено при изучении экспрессии брадизоитного антигена (Радка и др., 2005). Эти данные согласуются с наблюдениями, что спонтанная брадизоитная дифференциация происходит менее легко в быстро делящиеся штаммы T. гондия, такие как RH, и условие стресса, что медленный рост индуцирует дифференцировку брадизоитной (Bohne и др, 1994;. Иероним и др., 1998; Вайс и Ким, 2000).
Bradyzoites может пройти асинхронное разделение в вакуоли, в результате чего вакуолей с нечетным числом организмов вместо обычного кратных двум видели в тахизоит вакуолю (Dzierszinski и др., 2004). Брадизоитный дифференцировки не может быть отсоединен от замедления клеточного цикла, и может представлять собой стохастическое событие, которое происходит в определенной точке в клеточном цикле, когда репликация замедлилось достаточно. Оказывается, что все условия, которые медленно (но не арестовывать) результат клеточного цикла в брадизоитных дифференциациях, в то время как условия, которые блокируют клеточный цикл Прогрес-Sion не приводят к заметной дифференциации (Fox и Bzik., 2002; Хан и др, 2002).
При bradyzoites освобождаемый из цистой ткани используют для инфицирования клеток-хозяв, тахизоит специфический анти-род (например, SAG1) выражены в течение 15 часов после инфицирования до какого-либо значительного деления клеток путем вдувания брадизоитного (Soete и Dubremetz, 1996). К 48 ч пи, вакуоли, содержащие органно-измы, экспрессирующие только тахизоит антигены очевидны. Аналогичным образом, тахизоиты использовали для инфицирования с воздействием напряжений (рН 8,1) выражают брадизоитный антигены в течение 24 часов, и вакуоль, содержащий один или два организма может быть найдено, которые имеют брадизоитный антигены. Преобразование между этими стадиями развития, поэтому быстрое
событие, и стремление к дифференциации может происходить во время или вскоре после вторжения.
Хотя небольшая часть репликации пара-сайтов (<10 процентов) имеют от 1,8 до содержания 2ны ДНК (т.е. находятся в премитотическоге состояния G2), паразиты, которые одновременно экспрессируют брадизоитный маркер BAG1 и тахизоит маркер SAG1 имеют гораздо больше шансов иметь G2 премитотическое содержание ДНК - примерно 50 процентов от этих паразитов (Радка и др., 2003). Интересно, что проточная цитометрия измерение содержания ДНК зрелых bradyzoites, выделенные из тканевых кист демонстрирует, что этот этап имеет содержание 1N ДНК, в соответствии с их существами в состоянии покоя Gосостояние (Радка и др., 2003). В целом представляется, что приверженность брадизоитной дифференциации, вероятно, происходит в определенной точке клеточного цикла и что транзит через клеточный цикл необходим для дифференцировки. Этот ранние «предварительно bradyzoites» может продолжать репликацию, но в каком-то момент в развитии и созревании ткани кисты в полностью зрелом bradyzoites ввести покой Gо государство.
СТРЕСС РЕАКЦИЯ
И BRADYZOITES
В то время как дифференциация запрограммированный ответ, брадизоитная дифференцировка также связанные со стрессом ответ Т. гондия к условиям окружающей среды - например, воспалительный ответ хозяина. Много различного классический ответа индуцирующего стресса-условие, такие как температура, рН и митохондриальные ингибиторы, которые связаны с развитием брадизоитным в пробирке. Брадизоитная дифференциация, вероятно, разделяет общие черты с другими стрессом-индуцированной дифференциацией систем, такие как глюкоза голод и образование гифа грибов в спорообразовании или в Dicyostelium (Soderbom и Loomis, 1998; Томасон и др., 1999). Эти системы демонстрируют уникальные белки, связанные с конкретными дифференциациями струк-р в каждом организме, а также использование филогенетический законсервированные пути. Многие из этих сигнальных путей включают циклические нуклеотиды и киназы как часть системы регулирования в дифференциации. Существует значительное
| СТРЕСС РЕАКЦИЯ И BRADYZOITES |
корпус данных, касающихся белки теплового шока с дифференциацией в различных фил (Хейккиля, 1993a, 1993b).
BAG1 (также известный как BAG5) является одним из наиболее распространенных брадизоитных специфических генов, найденных в брадизоитном ЭЧТЕ Т. гондий, что составляет около 3 процентов всех брадизоитных специфических клонов. Этот ген имеет гомологию с малых белков теплового шока, и, следовательно, также называется hsp30 (Bohne и др., 1995;. Parmley и др, 1995). Оба BAG1 мРНК и белки (цитоплазматический антиген 28-кДа) являются повышались во время формирования брадизоитного, предполагая регуляцию транскрипции его Expres-Sion. Токсоплазма, экспрессирующие BAG1 рассматривается в течение 24 часов после контакта с рНом 8,0 или другими стрессовыми условиями.
Карбоксильной-концевой области BAG1 имеет небольшой тепловой шок мотив наиболее похожий на малые белки теплового шока растений, а рядом с N-конца является синапсины Iа-подобный домен, который может быть вовлечен в ассоциации этого небольшого тепла -ударных белка с белками в процессе разработки. Дрожжи дигибридная скрининг как с N-концевых и С-концевых доменов имеет Identi-Fied потенциал BAG1 взаимодействующих белков, которые в настоящее время подтверждены (Weiss и Ма, unpub-ликовано данные). BAG1 по-видимому, присутствуют, главным образом, в качестве мономера, и карбокси-концевой области BAG1 не реагируют с собой в дрожжевой двухгибридный скрининг (Weiss и Ма, неопубликованные данные). Четыре других мелких белки теплового шока присутствуют в Т. гондий; из них, hsp20, hsp21 и hsp29 выражаются в обеих тахизоитах и bradyzoites и hsp28 специфично для тахизоитов (де Мигель, 2005). Ни один из этих других малых белков теплового шока не связан с брадизоитным дифференциальным-ции, и все они присутствуют в виде мультимерных форм в цитозоле Т. гондий.
Гомолог белка теплового шока 70 (hsp70) индуцируется во время перехода как от тахи- zoite к брадизоитному и от брадизоитного к тахи- zoite (Lyons и Johnson, 1995, 1998; Silva и соавт., 1998; Weiss и соавт.., 1998; Миллер и др, 1999). Индукция hsp70 может быть продемонстрирована на обоих белков и уровней РНК. Кверцетин, ингибитор синтеза Hsp, может подавлять Hsp70 и уменьшить способность рН шок вызывать брадизоитный
Образование (Вайс и др., 1996, 1998). Внеклеточный Т. гондий обрабатывали 1-часовой экспозиции до рН 8,1 по сравнению с рН 7,1 выражают индуцируемый hsp70 72-кДа (обнаруженный с монАТ C92F3A-5; Stressgen) (Weiss. И др, 1998), и это внеклеточная Лечение вызывает образование брадизоитный. Т. гондий-инфицированные культуры, обработанные с рНом 8,1 показывает четыре раза induc-Тион уровней hsp70 по сравнению с T.gondii, выращенной в рНе 7,1-обработанные клетки (Weiss и соавт., 1996, 1998), которая аналогична величина ответа hsp70 продемонстрировали в Trypanosoma Тгурапозота, Theileria членистых и Plasmodium Falci-parum (Шилс и др., 1997). Подобное увеличение hsp70 видно цистами в естественных условиях при реактивации в мышиной модели индуцированного антиγинтерферон (Сильва и др., 1998). Относительный уровень экспрессии hsp70 Т. гондий также связан с Виру-Ленчик, и штамм RH имеет четыре копии повторяющегося звена семь-амино-кислоты (GGMPGGM) на С-конце его hsp70, по сравнению с пятью копиями в штамме ME49 (Lyons и Johnson, 1998).
В дополнение к hsp70, hsp90, уровни мРНК и белка также увеличивают во брадизоитный дифференциальными-ции (Эчеверрия и др., 2005). Флуоресцентная микроскопия показала, что в тахизоитах HSP90 находятся в цитозоле, тогда как в зрелом Brady-zoites hsp90 присутствует как в ядре и цитозоле. В Т. Gondii мутантов, которые не способны различать, hsp90 обнаруживается только в цитоплазме. Гелданамицин, A бензохинон ansamycin antibi-ушной способен связывать и нарушая функцию hsp90, блоки преобразования как из тахизоят в брадизоитных и брадизоитном к тахизоит (Эчеверрия и др., 2005).
Экспрессия репортерных генов под контролем промотора hsp70 также реагирует на условия, которые индуцируют образование брадизоитного (Мо и др., 2004). РН регулируется цис-элемент карты hsp70 промотора в регион-420 через -340 от исходного ATG гена hsp70 (Ma и др., 2004). В-650 пар оснований от исходного ATG представляет собой последовательность AGAGACG, которая была описана как
a цис- действующий элемент, который действует в качестве усилителя в транскрипции нескольких генов Т. гондий (Mercier
и другие., 1996). Существует ряд nGAAn повторов-385 от исходного ATG, которые имеют сходство с тепловой шок элемент (HSE), описанный в других
350 РАЗВИТИЕ брадизоитного
эукариоты (Morimot и др., 1994). CCGGGG находится рядом с этим HSE похож на sp1-hsp70 сайт в промоторе hsp70 человека (Morgan, 1989).
Кроме того, промотор HSP70 содержит несколько AGGGG или CCCCT области, которые являются Сими-лар в основной области (элемента стресс-ответа) Stre описаны во многих эукариотических генов (Эструч, 2000). Подобно STRE и HSE элементы рассматриваются в промоторной области енолазы 1, брадизоитный конкретной изоформы енолазы (Kibe и др., 2005). У дрожжей енолаза также известен как hsp48, так как это связано со стрессом белков теплового шока (Иида и Яхара, 1985). STRE-связывающая активность обнаружено в ядерном экстракте из индуцированного стресса bradyzoites значительно выше, чем от ненапряженных тахизоитов (Kibe и др., 2005). Факторы транскрипции, ответственные за регулирование Hsp70 и енолазов 1 еще не были определены, хотя анализы сдвига electromo-Билить (EMSA) предполагают, что существует специфические белки, которые распознают элементы Stre и HSE этих генов (Мо и др., 2004;. Kibe и др, 2005). Несмотря на то, есть область сходства между промоторной областью BAG1 и что из hsp70 области промотора, олигонуклеотиды из этого региона BAG1 вверх по течению, не конкурируют в EMSA (Мо и др., 2004). (См глава 16 для дальнейшего обсуждения регуляции экспрессии генов).
В эукариот, клеточная реакция на стресс является Asso-лем, связанных с фосфорилированием альфа-субъединицы eIF2 (эукариотической инициации фактор-2) повышение поступательную экспрессии BZIP белков, таких как GCN4 в дрожжах и ATF4 у млекопитающих (Хиннебуш и Natarajan, 2002). Роман eIF2 протеинкиназы, обозначенный TgIF2K-А (токсоплазма инициирования фактор-2-киназа) был идентифицирован и eIF2 фос-phorylation усиливается услови х, известных для индукции дифференцировки брадизоитный. Анализ генома Т. гондий демонстрирует гомологи генов, вовлеченных в этот стресс пути реакции, такие как GCN1 и GCN20 (Sullivan и др., 2004), а также гистона acetyltransferae GCN5 (Hettmann и Soldati, 1999; Sullivan и Смит, 2000) и гомолог SWI / SNF (Sullivan и др., 2003). Оба GCN5 и SWI / SNF, как известно, завербован GCN4 в дрожжах,
Эпигенетические события, такие как модификации гистонов, вероятно, являются ключевыми факторами в процессе дифференцировки, что приводит к образованию брадизоитного. TgSRCAP (токсоплазма SNF2 связанных с ОШП активатор протеина) является SWI2 / SNF2 семьи CHRO-Матин перемодельер, экспрессия которого увеличивается во время развития кист (Nallani и Sullivan, 2005). Как и в других организмах, ацетилирование гистонов и структуры метилирования коррелируют с геномом Acti-vation или репрессией в Т. гондиях (Saksouk и соавт, 2005). В брадизоитных дифференциациях доходов, маркера активации гена связаны с брадизоитными промоторами (Saksouk и соавт, 2005). Эпигенетический регулируемые изменения в экспрессии генов, а также изменения в хроматине обычно требуют модификаций транна-сидеть через фазу S, как уже были отмечена для индукции экспрессии маркеров брадизоитных (Радка и др., 2003). Геномные-широкий анализ эпигенетических изменений, которые происходят во время стадии отличаются друг-entiation продолжаются (Gissot и Ким, unpub-ликовано). Интересно отметить, что в Dictyostelium, GCN2 ортолог участвует в развитии плодового тела в ответ на стресс питательных веществ (Fang и др., 2003).