Образовавшиеся глицерол-3-фосфат и ацил-КоА жирных кислот – исходные соединения для биосинтеза жира (см. рис. 11, этап V).
На первом этапе синтеза триацилглицеролов образуется 3-фосфатидная кислота (диацилглицерол-3-фосфат), при этом ацильные остатки КоА-производных жирных кислот переносятся специфическими ацил-трансферазами на спиртовые группы глицерол-3-фосфата:

На втором этапе фосфатидная кислота гидролизуется фосфатидат-фосфогидролазой с образованием 1,2-диацилглицерола:
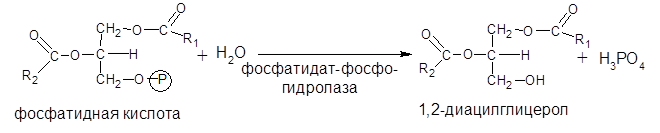
Затем 1,2-диацилглицерол превращается в триацилглицерол путем переноса остатка КоА-производного третьей жирной кислоты. Этот перенос катализируется ферментом диацилглицерол-ацилтрансферазой:
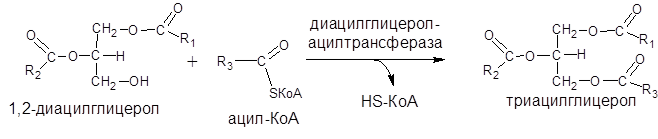
Метаболизм фосфолипидов
Распад (катаболизм) фосфолипидов
Фосфолипиды так же как и жиры, активно распадаются в тканях животных и растений.
Гидролиз фосфолипидов осуществляют несколько фосфолипаз, различающихся специфичностью, получивших название фосфолипаз А, С и Д:
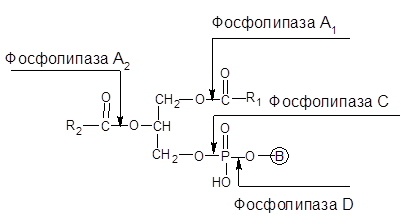
Здесь R1 и R2 – углеводородные радикалы; В – остатки азотсодержащих одноатомных спиртов (холина, этаноламина, серина).
Фосфолипаза А1 отщепляет жирные кислоты в α-положении; фосфолипаза А2 осуществляет ту же самую реакцию в β-положении; фосфолипаза С отщепляет фосфорилированные азотсодержащие спирты от фосфолипидов; фосфолипаза D гидролизует фосфолипиды с образованием азотсодержащего спирта и фосфатидной кислоты.
В растениях обнаружены все виды фосфолипаз. Фосфолипазы А обнаружены также у животных и микроорганизмов. Фосфолипазы локализованы преимущественно в лизосомах. Конечными продуктами гидролиза фосфолипидов являяются глицерин, жирные кислоты, азотистые спирты и фосфорная кислота.
Синтез фосфолипидов
В отличие от триацилглицеролов и жирных кислот фосфолипиды не являются существенным энергетическим материалом. Фосфолипиды, как было сказано ранее, играют важную роль в структуре и функции клеточных мембран.
Фосфолипиды синтезируются, главным образом, в эндоплазматической сети клетки.
Центральную роль в биосинтезе фосфолипидов играют L-диацилглицеролы (в синтезе фосфатидилхолинов и фосфатидилэтаноламинов), фосфатидная кислота (в синтезе фосфатидилинозитов) и сфингозин (в синтезе сфингомиелинов).
В этих биосинтезах принимает участие цитидинтрифосфат (ЦТФ), выполняющий роль активатора – поставщика энергии.
Основными компонентами мембран являются фосфатидилэтаноламин и фосфатидилхолин.
В процессе синтеза фосфатидилэтаноламина вначале происходит активация этаноламина под действием этаноламинкиназы:

Этаноламинфосфат реагирует затем с цитидинтрифосфатом (ЦТФ), в результате чего образуется цитидиндифосфатэтаноламин (ЦДФ-этаноламин) и пирофосфат (РРі). Эту реакцию катализирует фосфоэтаноламинцитидинтрансфераза:
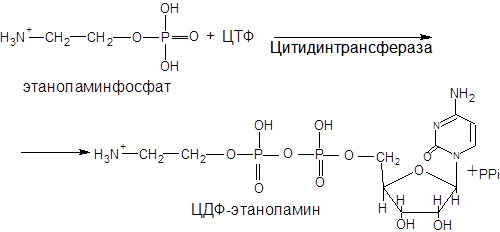
Активированный ЦДФ–этаноламин при участии фермента этаноламинфосфаттрансферазы вступает в реакцию с L–диацилглицеролом с образованием фосфатидиламина:

L-диацилглицерол образуется при гидролизе фосфатидной кислоты. Аналогичные реакции ведут к образованию ЦДФ-холина.; последний вступает в реакцию с L-диацилглицеролом, в результате чего образуется фосфатидилхолин.
В синтезе фосфатидилэтаноламина и фосфатидилхолина важную роль играет ЦТФ.
Фосфолипиды в клетке легко подвергаются взаимопревращениям. Фосфатидилэтаноламин является предшественником фосфатидилхолина. Так, в печени в результате последовательного переноса трех метильных групп от трех молекул S-аденозилметионина образуется фосфатидилхолин:
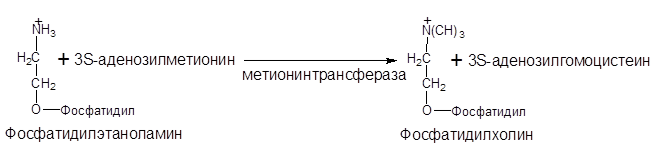
В переносе метильных групп участвуют тетрагидрофолевая кислота (витамин Вс) и метилкобаламин (витамин В12).
Синтез фосфатидилсерина происходит другим путем с предварительным вовлечением фосфатидной кислоты:

Затем происходит перенос серина на фосфатидильный остаток с образованием фосфатидилсерина:
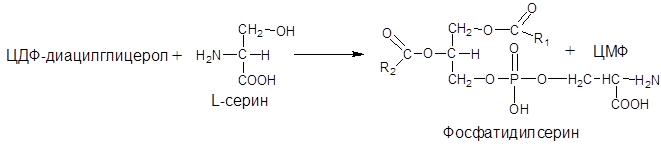
У млекопитающих фосфатидилсерин образуется в реакции обмена этаноламина на серин следующим путем:
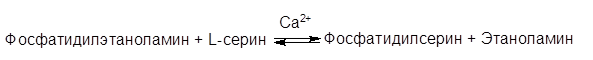
Таким же путем образуется фосфатидилинозитол.
В биосинтезе сфингомиелина принимает участие церамид (N-ацетилсфингозин), который образуется при взаимодействии спирта сфингозина и ацетил-КоА. Сфингомиелин синтезируется в результате взаимодействия церамида с ЦДФ-холином:

Все выше синтезированные фосфолипиды поступают с помощью липидпереносящих белков из цитоплазмы к мембранам и участвуют в их построении.
Список используемой литературы
1. Березов Т.Т. Биологическая химия: Учебник для вузов/ Т.Т. Березов, Б.Ф.Коровкин. –3-е изд., перераб. и доп. – М.: Медицина, 1998. – 704 с.
2. Филиппович Ю.Б. Основы биохимии. – М.: Ангар, 1999. – 512 с
3. Жеребцов Н.А., Попова Т.Н., Артюхов В.Г. Биохимия: Учебник. – Воронеж: Из-во ВГУ, 2002. – 696 с.
4. Биохимия растительного сырья. /Под. ред. В.Г. Щербакова.– М.: Колос, 1999. – 376 с.
5. Ленинджер А. Биохимия. – М.: Мир, 1999. Т1-3.
6. Основы биохимии. /Под ред. Анисимова. – М.: Высшая школа, 1986. – 551 с.
7. Кретович В.Л. Биохимия растений. – М.: Высшая школа, 1986. – 503 с.
8. Взаимосвязь процессов обмена углеводов, жиров и белков. Методические указания по дисциплине «Биологическая химия» для студентов 49 01 01, 49 01 02, 91 01 01, 48 01 02.– Могилев: МГУП, 2005. – 43 с.
КОНСПЕКТ ЛЕКЦИЙ
БИОЛОГИЧЕСКАЯ ХИМИЯ
Методические указания
по дисциплине «Биологическая химия»
для студентов специальностей
1-49 01 01, 1-49 01 02, 1-91 01 01
Составители: Ольга Николаевна Макасеева
Лариса Михайловна Ткаченко
Редактор Татьяна Леонидовна Бажанова
Технический редактор Анна Аполлинариевна Щербакова
Подписано в печать_______________ Формат 60х84 1/16
Печать офсетная. Усл. печ. л. ____________ Уч.-изд.л.__________
Тираж________экз. Заказ__________ Бесплатно
ЛП № 226 от 12.02.2003 г.
ЛИ № 604 от 03.06.2003г.
Отпечатано на ризографе УО МГУП
212007, Могилев, пр-т Шмидта, 3