Желудок является отделом пищеварительного тракта, в котором пища, смешанная со слюной, покрытая вязкой слизью слюнных желез пищевода, задерживается от 3 до 10 часов для ее механической и химической обработки. К функциям желудка относятся: (1) депонирование пищи; (2) секреторная — отделение желудочного сока, обеспечивающего химическую обработку пищи; (3) — двигательная — перемешивание пищи с пищеварительными соками и ее передвижение порциями в двенадцатиперстную кишку; (4) — всасывания в кровь незначительных количеств веществ, поступивших с пищей. Вещества, растворенные в спирту, всасываются в значительно больших количествах; (5) — экскреторная — выделение вместе с желудочным соком в полость желудка метаболитов (мочевина, мочевая кислота, креатин, креатинин), концентрация которых здесь превышает пороговые величины, и веществ, поступивших в организм извне (соли тяжелых металлов, йод, фармакологические препараты); (6) — инкреторная — образование активных веществ (гормонов), принимающих участие в регуляции деятельности желудочных и других пищеварительных желез (гастрин, гистамин, соматостатин, мотилин и др.); (7) — защитная — бактерицидное и бактериостатическое действие желудочного сока и возврат недоброкачественной пищи, предупреждающий ее попадание в кишечник.
Секреторная деятельность желудка осуществляется желудочными железами, продуцирующими желудочный сок и представленными тремя видами клеток: главными (главные гландулоциты), принимающие участие в выработке ферментов; париетальными (париетальные гландулоциты), участвующие в выработке хлористоводородной кислоты (НС1) и добавочными (мукоциты), выделяющими мукоидный секрет (слизь).
Клеточный состав желез изменяется в зависимости от принадлежности их к тому или иному отделу желудка, соответственно изменяется состав и свойства секрета, который они выделяют.
Состав и свойства желудочного сока. В состоянии покоя "натощак" из желудка человека можно извлечь около 50 мл желудочного содержимого нейтральной или слабокислой реакции (рН=б,0). Это смесь слюны, желудочного сока (так называемая "базальная" секреция), а иногда — забрасываемое в желудок содержимое двенадцатиперстной кишки.
Общее количество желудочного сока, отделяющегося у человека при обычном пищевом режиме, составляет 1,5-2,5 л в сутки. Это
398
бесцветная, прозрачная, слегка опалесцируюшая жидкость с удельным весом 1,002-1,007. В соке могут быть хлопья слизи. Желудочный сок имеет кислую реакцию (рН=0,8-1,5) вследствие высокого содержания в нем хлористоводородной кислоты (0,3-0,5%). Содержание воды в соке 99,0-99,5% и 1,0-0,5% — плотных веществ. Плотный остаток представлен органическими и неорганическими веществами (хлоридами, сульфатами, фосфатами, бикарбонатами натрия, калия, кальция, магния). Основной неорганический компонент желудочного сока — хлористоводородная кислота — может быть в свободном и связанном с протеинами состоянии. Органическая часть плотного остатка — это ферменты, мукоиды (желудочная слизь), один из них — гастромукопротеид (внутренний фактор Кастла), необходим для всасывания витамина В12. В небольшом количестве здесь находятся азотсодержащие вещества небелковой природы (мочевина, мочевая кислота, молочная кислота и др.).
Рис.9.2. Образование соляной кислоты желудочного сока. Пояснения в тексте.
Механизм секреции хлористоводородной кислоты. Хлористоводородная кислота (НС1) вырабатывается париетальными клетками, расположенными в перешейке, шейке и верхнем отделе тела железы (рис.9.2). Эти клетки характеризуются исключительным богатством митохондрий вдоль внутриклеточных канальцев. Площадь мембраны

399
канальцев и апикальной поверхности клеток невелика и при отсутствии специфической стимуляции в цитоплазме этой зоны имеется большое количество тубовезикул. Во время стимуляции на высоте секреции создается избыток площади мембран в результате встроенных в них тубовезикул, что сопровождается значительным увеличением клеточных канальцев, проникающих вплоть до базальной мембраны. Вдоль вновь образованных канальцев располагается множество четко структурированных митохондрий, площадь внутренней мембраны которых возрастает в процессе биосинтеза НС1. Число и протяженность микроворсинок многократно возрастает, соответственно увеличивается площадь контакта канальцев и апикальной мембраны клетки с внутренним пространством железы. Увеличение площади секреторных мембран способствует наращиванию в них числа ионных переносчиков. Таким образом, увеличение секреторной активности париетальных клеток обусловливается увеличением площади секреторной мембраны. Это сопровождается повышением суммарного заряда ионного переноса, и увеличением числа контактов мембран с митохондриями — поставщиками энергии и ионов водорода для синтеза НС1.
Кислопродуцирующие (оксинтные) клетки желудка активно используют собственный гликоген для нужд секреторного процесса. Секреция НС1 характеризуется как ярко выраженный цАМФ-зависимый процесс, активация которого протекает на фоне усиления гликоге-нолитической и гликолитической активности, что сопровождается продукцией пирувата. Окислительное декарбоксилирование пирувата до ацетил-КоА-СО2 осуществляется пируватдегидрогеназным комплексом и сопровождается накоплением в цитоплазме НАДН2. Последний используется для генерирования Н+ в процессе секреции НС1. Расщепление триглицеридов в слизистой желудка под влиянием триглицеридлипазы и последующая утилизация жирных кислот создает в 3-4 раза больший приток восстановительных эквивалентов в митохондриальную цепь переноса электронов. Обе цепи реакции, как аэробный гликолиз, так и окисление жирных кислот, запускаются посредством цАМФ-зависимого фосфорилирования соответствующих ферментов, обеспечивающих генерирование ацетил- КОа в цикле Кребса и восстановительных эквивалентов для электронпере-носящей цепи митохондрий. Са2+ выступает здесь как абсолютно необходимый элемент секреторной системы НС1.
Процесс цАМФ- зависимого фосфорилирования обеспечивает активацию желудочной карбангидразы, роль которой как регулятора кислотно-щелочного равновесия в кислотопродуцирующих клетках особенно велика. Работа этих клеток сопровождается длительной и массовой потерей ионов Н+ и накоплением в клетке ОН, способных оказать повреждающее действие на клеточные структуры. Нейтрализация гидроксильных ионов и является главной функцией карбангидразы. Образующиеся бикарбонатные ионы посредством электронейтрального механизма выводятся в кровь, а ионы CV входят в клетку.
Кислотопродуцирующие клетки на наружных мембранах имеют две мембранные системы, участвующие в механизмах продукции Н+ и
400
секреции НС1 — это Na+, К+-АТФаза и (Н++К+)-АТФаза. Na+, K+-АТФаза, расположенная в базолатеральных мембранах, переносит К+ в обмен на Na+ из крови, а (Н++К+)-АТФаза, локализованная в секреторной мембране, транспортирует калий из первичного секрета в обмен на выводимые в желудочный сок ионы Н+.
В период секреции митохондрии всей своей массой в виде муфты, охватывают секреторные канальцы и их мембраны сливаются, образуя митохондриалъно-секреторный комплекс, где ионы Н+ могут непосредственно акцентироваться (Н++К+)-АТФазой секреторной мембраны и транспортироваться из клетки.
Таким образом, кислотообразующая функция обкладочных клеток характеризуется наличием в них процессов фосфорилирования — дефосфорилирования, существованием митохондриальной окислительной цепи, транспортирующей ионы Н+ из матриксного пространства, а также (Н++К+)-АТФазы секреторной мембраны, перекачивающей протоны из клетки в просвет железы за счет энергии АТФ.
Вода поступает в канальцы клетки путем осмоса. Конечный секрет, поступающий в канальцы, содержит НС1 в концентрации 155 ' ммоль/л, хлористый калий в концентрации 15 ммоль/л и очень малое количество хлористого натрия.
Роль хлористоводородной кислоты в пищеварении. В полости желудка хлористоводородная кислота (НС1) стимулирует секреторную активность желез желудка; способствует превращению пепсиногена в пепсин, путем отщепления ингибирующего белкового комплекса; создает оптимальное рН для действия протеолитических ферментов желудочного сока; вызывает денатурацию и набухание белков, что способствует их расщеплению ферментами; обеспечивает антибактериальное действие секрета. Хлористоводородная вода способствует также переходу пищи из желудка в двенадцатиперстную кишку; участвует в регуляции секреции желудочных и поджелудочных желез, стимулируя образование гастроинтестинальных гормонов (гастрина, секретина); стимулирует секрецию фермента энтерокиназы энтеро-цитами слизистой двенадцатиперстной кишки; участвует в створаживании молока, создавая оптимальные условия среды и стимулирует моторную активность желудка.
Помимо хлористоводородной кислоты в желудочном соке в небольших количествах содержатся кислые соединения — кислые фосфаты, молочная и угольная кислоты, аминокислоты.
Ферменты желудочного сока. Основным ферментативным процессом в полости желудка является начальный гидролиз белков до альбумоз и пептинов с образованием небольшого количества аминокислот. Желудочный сок обладает протеолитической активностью в широком диапазоне рН с оптимумом действия при рН 1,5-2,0 и 3,2-4,0.
В желудочном соке выделено семь видов пепсиногенов, объединенных общим названием пепсины. Образование пепсинов осуществляется из неактивных предшественников — пепсиногенов, находя-
401
щихся в клетках желудочных желез в виде гранул зимогена. В просвете желудка пепсиноген активируется НС1 путем отщепления от него ингибирующего белкового комплекса. В дальнейшем, в ходе секреции желудочного сока активация пепсиногена осуществляется аутокаталитически под действием уже образовавшегося пепсина.
При оптимальной активности среды песин оказывает лизирующее действие на белки, разрывая в белковой молекуле пептидные связи, образованные группами фениламина, тирозина, триптофана и других аминокислот. В результате этого воздействия белковая молекула распадается на пептоны, протеазы и пептиды. Пепсин обеспечивает гидролиз главных белковых веществ, особенно коллагена — основного компонента волокон соединительной ткани.
Основными пепсинами желудочного сока являются:
- пепсин А — группа ферментов, гидролизирующих белки при рН=1,5-2,0. Часть пепсина (около 1%) переходит в кровеносное русло, откуда вследствие небольшого размера молекулы фермента проходит через клубочковый фильтр и выделяется с мочой (уропеп- син). Определение содержания уропепсина в моче используется в лабораторной практике для характеристики протеолитической актив ности желудочного сока;
- гастриксин, пепсин С, желудочный катепсин — оптимум рН для ферментов этой группы является 3,2-3,5. Соотношение между пепсином А и гастриксином в желудочном соке человека от 1:1 до 1:5;
- пепсин В, парапепсин, желатиназа — разжижает желатину, расщепляет белки соединительной ткани. При рН—5,6 и выше дей ствие фермента угнетается;
- реннин, пепсин Д, химозин — расщепляют казеин молока в присутствии ионов Са++, с образованием параказеина и сывороточ ного белка.
Содержание пепсинов и гастриксина в слизистой различных отделов желудка неодинаково: пепсины отсутствуют в антральном отделе желудка, гастриксин же присутствует во всех отделах желудка.
Желудочный сок содержит ряд непротеолитических ферментов. Это — желудочная липаза, расщепляющая жиры, которые находятся в пище в эмульгированном состоянии (жиры молока), на глицерин и жирные кислоты при рН=5,9-7,9. У детей желудочная липаза расщепляет до 59% жира молока. В желудочном соке взрослых людей липазы мало. Лизоцим (мурамидаза), имеющийся в желудочном соке, обладает антибактериальным действием. Уреаза — расщепляет мочевину при рН=8,0. Освобождающийся при этом аммиак нейтрализи-рует НС1.
Желудочная слизь и ее роль в пищеварении. Обязательным органическим компонентом желудочного сока является слизь, которая продуцируется всеми клетками слизистой оболочки желудка. Наибольшую мукоидпродуцирующую активность проявляют добавочные клетки (мукоциты). В состав слизи входят нейтральные мукополи-сахариды, сиаломуцины, гликопротеины и гликаны.
Нерастворимая слизь (муцин) является продуктом секреторной активности добавочных клеток (мукоциты) и клеток поверхностного эпителия желудочных желез. Муцин освобождается через апикальную мембрану, образует слой слизи, обволакивающий слизистую оболочку желудка и препятствующий повреждающим воздействиям экзогенных факторов. Этими же клетками одновременно с муцином продуцируется бикарбонат. Образующийся при взаимодействии муцина и бикарбоната мукозо-бикарбонатный барьер предохраняет слизистую от аутолиза под воздействием соляной кислоты и пепсинов.
При рН ниже 5,0 вязкость слизи уменьшается, она растворяется и удаляется с поверхности слизистой оболочки, при этом в желудочном соке появляются хлопья, комочки слизи. Одновременно со слизью удаляется адсорбированные ею ионы водорода и протеиназы. Таким образом формируется не только механизм защиты слизистой оболочки, но и происходит активация пищеварения в полости желудка.
Нейтральные мукополисахариды (основная часть нерастворимой и растворимой слизи) являются составной частью групповых антигенов крови, фактора роста и антианемического фактора Кастла.
Сиаломуцины, входящие в состав слизи, способны нейтрализовать вирусы и препятствовать вирусной гемаглютинации. Они же участвуют в синтезе НС1.
Гликопротеины, вырабатываемые париетальными клетками, являются внутренним фактором Кастла, необходимым для всасывания витамина В,,. Отсутствие этого фактора приводит к развитию заболевания, известного под названием В12-дефицитной анемии (желе-зодефицитная анемия).
Регуляция желудочной секреции. В регуляции секреторной деятельности желудочных желез участвуют нервный и гуморальный механизмы. Весь процесс желудочного сокоотделения условно можно разделить на три наслаивающиеся друг на друга во времени фазы: сложнорефлекторную (цефалическую), желудочную и кишечную.
Первоначальное возбуждение желудочных желез (первая цефали-ческая или сложнорефлекторная фаза) обусловлено раздражением зрительных, обонятельных и слуховых рецепторов видом и запахом пищи, восприятием всей обстановки, связанной с приемом пищи (условнорефлекторный компонент фазы). На эти воздействия наслаиваются раздражения рецепторов ротовой полости, глотки, пищевода при попадании пищи в ротовую полость, в процессе ее жевания и глотания (безусловнорефлекторный компонент фазы).
Первый компонент фазы начинается с выделения желудочного сока в результате синтеза афферентных зрительных, слуховых и обонятельных раздражений в таламусе, гипоталамусе, лимбической системе и коре больших полушарий головного мозга. Это создает условия для повышения возбудимости нейронов пищеварительного бульбарного центра и запуска секреторной активности желудочных желез.
Раздражение рецепторов ротовой полости, глотки и пищевода передается по афферентным волокнам V, IX, X пар черепномозго-вых нервов в центр желудочного сокоотделения в продолговатом
403


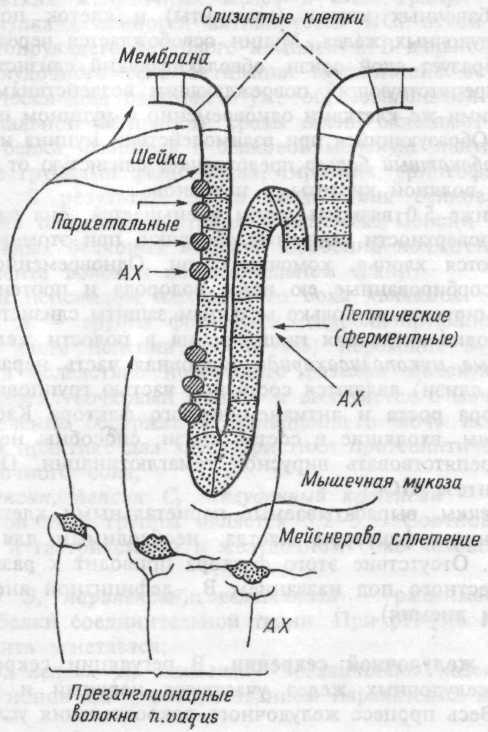
Рис.9.3. Нервная регуляция желудочных желез.
м озге. От центра импульсы по эфферентным волокнам блуждающегонерва направляются к желудочным железам, что приводит к дополнительному безусловнорефлекторному усилению секреции (рис.9.3). Сок, выделяющийся под влиянием вида и запаха пищи, жевания и глотания, получил название "аппетитного" или запального. Вследствие его выделения желудок оказывается заранее подготовленным к приему пищи. Наличие этой фазы секреции было доказано И.П.Павловым в классическом эксперименте с мнимым кормлением у эзо-фаготомированных собак.
Желудочный сок, полученный в первую сложнорефлекторную фазу, обладает высокой кислотностью и большой протеолитической активностью. Секреция в эту фазу зависит от возбудимости пищевого центра, легко тормозится при воздействии различных внешних и внутренних раздражителей.
404
На первую сложнорефлекторную фазу желудочной секреции наслаивается вторая — желудочная (нейрогуморалъная). В регуляции желудочной фазы секреции принимают участие блуждающий нерв, местные интрамуральные рефлексы. Выделение сока в эту фазу связано с рефлекторным ответом при действии на слизистую оболочку желудка механических и химических раздражителей (пища, попавшая в желудок, соляная кислота, выделившаяся с "запальным соком", растворенные в воде соли, экстрактивные вещества мяса и овощей, продукты переваривания белков), а также стимуляцией секреторных клеток тканевыми гормонами (гастрин, гастамин, бомбезин).
Раздражение рецепторов слизистой оболочки желудка вызывает поток афферентных импульсов к нейронам стволового отдела мозга, что сопровождается увеличением тонуса ядер блуждающего нерва и значительным усилением потока эфферентных импульсов по блуждающему нерву к секреторным клеткам. Выделение из нервных окончаний ацетилхолина не только стимулирует деятельность главных и обкладочных клеток, но и вызывает выделение гастрина G-клетками антрального отдела желудка. Гастрин — наиболее сильный из известных стимуляторов обкладочных и в меньшей степени главных клеток. Кроме того, гастрин стимулирует пролиферацию клеток слизистой и увеличивает кровоток в ней. Выделение гастрина усиливается в присутствии аминокислот, дипептидов, а также при умеренном растяжении антрального отдела желудка. Это вызывает возбуждение сенсорного звена периферической рефлекторной дуги эн-теральной системы и через интернейроны стимулирует активность G-клеток. Наряду со стимуляцией обкладочных, главных и G-клеток, ацетилхолин усиливает активность гистидиндекарбоксилазы ECL- клеток, что приводит к повышению содержания гистамина в слизистой оболочке желудка. Последний выполняет роль ключевого стимулятора выработки соляной кислоты. Гистамин действует на Н2-рецепторы обкладочных клеток, он необходим для секреторной активности этих клеток. Гистамин оказывает также стимулирующее действие на секрецию желудочных протеиназ, однако, чувствительность зимогеновых клеток к нему невелика в связи с низкой плотностью Н2-рецепторов на мембране главных клеток.
Третья (кишечная) фаза желудочной секреции возникает при переходе пищи из желудка в кишечник. Количество желудочного сока, выделяющегося в эту фазу, не превышает 10% от общего объема желудочного секрета. Желудочная секреция в начальном периоде фазы возрастает, а затем начинает снижаться.
Увеличение секрета обусловлено значительным усилением потока афферентных импульсов от механо- и хеморецепторов слизистой 12-перстной кишки при поступлении из желудка слабокислой пищи и выделением гастрина G-клетками двенадцатиперстной кишки. По мере поступления кислого химуса и снижения рН дуоденального содержимого ниже 4,0 секреция желудочного сока начинает угнетаться. Дальнейшее угнетение секреции вызвано появлением в слизистой 12-ти перстной кишки секретина, который является антагонистом гастрина, но в то же время усиливает синтез пепсиногенов.
405
По мере наполнения 12-ти перстной кишки и увеличения концентрации продуктов белкового и жирового гидролиза угнетение секреторной активности нарастает под влиянием пептидов, выделяемых желудочно-кишечными эндокринными железами (соматостатин, ва-зоактивный кишечный пептид, холесцитокинин, желудочный инги-биторный гормон, глюкагон). Возбуждение афферентных нервных путей возникает при раздражении хемо- и осморецепторов кишечника поступившими из желудка пищевыми веществами.
Гормон энтерогастрин, образующийся в слизистой оболочке кишечника, является одним из стимуляторов желудочной секреции и в третьей фазе. Продукты переваривания пищи (особенно белки), всосавшись в кишечнике в кровь, могут стимулировать желудочные железы путем усиления образования гистамина и гастрина.
Стимуляция желудочной секреции. Часть нервных импульсов, возбуждающих желудочную секрецию, берет начало в дорзальных ядрах блуждающего нерва (в продолговатом мозге), достигает по его волокнам энтеральной системы, а затем поступает к желудочным железам. Другая часть секреторных сигналов возникает внутри самой энтеральной нервной системы. Таким образом, в нервной стимуляции желудочных желез принимают участие как центральная нервная система, так и энтеральная нервная система. Рефлекторные влияния поступают к желудочным железам по рефлекторным дугам двух видов. Первые — длинные рефлекторные дуги — включают структуры, по которым афферентные импульсы направляются от слизистой оболочки желудка к соответствующим центрам головного мозга (в продолговатый мозг, гипоталамус), эфферентные — направляются обратно к желудку по блуждающим нервам. Вторые — короткие рефлекторные дуги — обеспечивают осуществление рефлексов в пределах местной энтеральной системы. Стимулы, вызывающие возникновение этих рефлексов, возникают при растяжении стенки желудка, тактильных и химических (HCI, пепсин и др.) воздействиях на рецепторы слизистой оболочки желудка.
Нервные сигналы, поступающие к желудочным железам по рефлекторным дугам, стимулируют секреторные клетки и одновременно активируют G-клетки, продуцирующие гастрин. Гастрин представляет собой полипептид, секретируемый в двух формах: "большей гастрин", содержащий 34 аминокислоты (G-34), и меньшая форма (G- 17), в состав которой входят 17 аминокислот. Последний более эффективен.
Гастрин, поступающий к железистым клеткам с током крови, возбуждает париетальные клетки и в меньшей степени — главные. Скорость секреции соляной кислоты под влиянием гастрина может возрасти в 8 раз. Выделившаяся соляная кислота, в свою очередь, возбуждая хеморецепторы слизистой оболочки, способствует секреции желудочного сока.
Активация блуждающего нерва сопровождается также усилением активности гистидиндекарбоксилазы в желудке, вследствие чего в его слизистой оболочке увеличивается содержание гистамина. Пос-
406
ледний непосредственно действует на париетальные гландулоциты, значительно увеличивая секрецию НС1.
Таким образом, адетилхолин, освобождающийся на нервных окончаниях блуждающего нерва, гастрин и гистамин оказывают одновременно стимулирующее воздействие на желудочные железы, обусловливая выделение хлористоводородной кислоты. Секреция пепсинoгe -на главными гландулоцитами регулируется ацетилхолином (освобождающимся на окончаниях блуждающего нерва и других энтеральных нервов), а также воздействием хлористоводородной кислоты. Последнее связано с возникновением энтеральных рефлексов при раздражении НС1 рецепторов слизистой оболочки желудка, а также с выделением под влиянием НС1 гастрина, оказывающего прямое воздействие на главные гландулоциты.
Пищевые вещества и желудочная секреция. Адекватными возбудителями желудочной секреции являются вещества, употребляемые в пищу. Функциональные приспособления желудочных желез к различной пище выражаются в различном характере секреторной реакции на них желудка. Индивидуальная адаптация секреторного аппарата желудка к характеру пищи обусловлена ее качеством, количеством, режимом питания. Классическим примером приспособительных реакций желудочных желез являются изученные И.П.Павловым секреторные реакции в ответ на прием пищи, содержащей преимущественно углеводы (хлеб), белки (мясо), жиры (молоко).
Наиболее эффективным возбудителем секреции является белковая пища (рис.9.4). Белки и продукты их переваривания обладают выраженным сокогонным действием. После приема мяса развивается

Рис.9.4. Выделение желудочного и поджелудочного сока на различные пищевые вещества.
Желудочный сок — пунктирная линия, поджелудочный сок — сплошная линия.
407
довольно энергичная секреция желудочного сока с максимумом на 2-м часе. Длительная мясная диета приводит к усилению желудочной секреции на все пищевые раздражители, повышению кислотности и переваривающей силы желудочного сока.
Углеводная пища (хлеб) — самый слабый возбудитель секреции. Хлеб беден химическими возбудителями секреции, поэтому после его приема развивается ответная секреторная реакция с максимумом на 1-м часе (рефлекторное отделение сока), а затем резко уменьшается и на невысоком уровне удерживается продолжительное время. При длительном нахождении человека на углеводном режиме кислотность и переваривающая сила сока снижаются.
Действие жиров молока на желудочную секрецию осуществляется в две стадии: тормозную и возбуждающую. Этим объясняется тот факт, что после приема пищи максимальная секреторная реакция развивается только к концу 3-го часа. В результате длительного питания жирной пищей происходит усиление желудочной секреции на пищевые раздражители за счет второй половины секреторного периода. Переваривающая сила сока при использовании в пище жиров ниже по сравнению с соком, выделяющимся при мясном режиме, но выше, чем при питании углеводной пищей.
Количество отделяющегося желудочного сока, его кислотность, протеолитическая активность зависят также от количества и консистенции пищи. По мере увеличения объема пищи секреция желудочного сока возрастает.
Эвакуация пищи из желудка в двенадцатиперстную кишку сопровождается торможение желудочной секреции. Как и возбуждение, этот процесс по механизму действия является нейрогуморальным. Рефлекторный компонент этой реакции вызывается снижением потока афферентных импульсов от слизистой желудка, в значительно меньшей степени раздражаемой жидкой пищевой кашицей с рН выше 5,0, нарастанием потока афферентных импульсов от слизистой 12-ти перстной кишки (энтерогастральный рефлекс).
Изменения химического состава пищи, поступление продуктов ее переваривания в 12-ти перстную кишку стимулируют выделение из нервных окончаний и эндокринных клеток пилорического отдела желудка, 12-ти перстной кишки и поджелудочной железы пептидов (со-матостатина, секретина, нейротензина, ГИП, глюкагона, холецистоки-нина), что вызывает торможение продукции соляной кислоты, а затем желудочной секреции в целом. Тормозное влияние на секрецию главных и обкладочных клеток оказывают также простагландина группы Е.
Немаловажную роль в секреторной деятельности желудочных желез играют эмоциональное состояние человека и стресс. Среди непищевых факторов, усиливающих секреторную активность желудочных желез, наибольшее значение имеют стресс, раздражение и ярость, угнетающее тормозное влияние на активность желез оказывают страх, тоска, депрессивные состояния человека.
Длительные наблюдения за деятельностью секреторного аппарата желудка у человека позволили обнаружить выделение желудочного сока и в межпищеварительный период. В этом случае эффективны-
408
ми оказались раздражители, связанные с приемом пищи (обстановка, в которой обычно происходит прием пищи), заглатыванием слюны, забрасыванием в желудок дуоденальных соков (панкреатического, кишечного, желчи).
Плохо пережеванная пища или накапливающийся углекислый газ вызывает раздражение механо- и хеморецепторов слизистой оболочки желудка, что сопровождается активацией секреторного аппарата слизистой желудка и секрецией пепсинов и соляной кислоты.
Спонтанную секрецию желудка могут вызывать расчесы на коже, ожоги, абсцессы, она возникает у хирургических больных в послеоперационный период. Это явление связано с усиленным образованием гистамина из продуктов тканевого распада, его высвобождением из тканей. С током крови гистамин достигает желудочных желез и стимулирует их секрецию.
Моторная деятельность желудка. Желудок хранит, согревает, смешивает, размельчает, приводит в полужидкое состояние, сортирует и продвигает по направлению к 12-перстной кишке содержимое с различной скоростью и силой. Все это совершается благодаря двигательной функции, обусловленной сокращением его гладкомы-шечной стенки. Характерными свойствами ее клеток, как и мышечной стенки всей пищеварительной трубки, являются способности к спонтанной активности (автоматии), в ответ на растяжение — сокращаться и находиться в сокращенном состоянии длительное время. Мускулатура желудка может не только сокращаться, но и активно расслабляться.
Вне фазы пищеварения желудок находится в спавшем состоянии, без широкой полости между его стенками. Через 45-90 минут периода покоя возникают периодические сокращения желудка, длящиеся 20-50 минут (голодная периодическая деятельность). При наполнении пищей он приобретает форму мешка, одна сторона которого переходит в конус.
Во время приема пищи и спустя некоторое время стенка дна желудка расслаблена, что создает условия для изменения объема без значительного повышения давления в его полости. Расслабление мускулатуры дна желудка во время еды получило название "рецептивного расслабления".
В наполненном пищей желудке отмечены три вида движений: (1) перистальтические волны; (2) сокращение терминальной части мускулатуры пилорического отдела желудка; (3) уменьшение объема полости дна желудка и его тела.
Перистальтические волны возникают в течение первого часа после еды на малой кривизне вблизи пищевода (где находится карди-альный водитель ритма) и распространяются к пилорическому отделу со скоростью 1 см/с, длятся 1,5 с и охватывают 1-2 см желудочной стенки. В пилорическом отделе желудка длительность волны составляет 4-6 в минуту и ее скорость возрастает до 3-4 см/с.
Благодаря большой пластичности мышц стенки желудка и способности повышать тонус при растяжении пищевой комок, поступив-
409
ший в его полость, плотно охватывается стенками желудка, вследствие чего в области дна по мере поступления пищи образуются "слои". Жидкость стекает в антральный отдел независимо от величины наполнения желудка.
Если прием пищи совпадает с периодом покоя, то сразу же после еды возникают сокращения желудка, если же поступление пищи совпадает с голодной периодической деятельностью, то сокращения желудка тормозятся и возникают несколько позже (3-10 мин). В начальный период сокращений возникают мелкие низкоамплитудные волны, способствующие поверхностному смешиванию пищи с желудочным соком и перемещению небольших ее порций в тело желудка. Благодаря этому внутри пищевого комка продолжается расщепление углеводов амилолитическими ферментами слюны.
Редкие низкоамплитудные сокращения начального периода пищеварения сменяются более сильными и частыми, что создает условия для активного перемешивания и перемещения содержимого желудка. Однако пища продвигается вперед медленно, потому что волна сокращения проходит над комком пищи, увлекая его за собой, а затем отбрасывает его обратно. Таким образом, совершается механическая работа по размельчению пищи и ее химическая обработка благодаря многократному движению вдоль активной поверхности слизистой, насыщенной ферментами и кислым соком.
Перистальтические волны в теле желудка перемещают по направлению к пилорическому отделу часть пищи, подвергшейся воздействию желудочного сока. Эта порция пищи замещается пищевой массой из более глубоких слоев, что обеспечивает ее смешивание с желудочным соком. Несмотря на то, что перистальтическая волна формируется единым гладкомышечным аппаратом желудка, приближаясь к антральному отделу она утрачивает свой плавный поступательный ход и происходит тоническое сокращение антрального отдела.
В пилорическом отделе желудка возникают пропульсивные сокращения, обеспечивающие эвакуацию содержимого желудка в 12-ти перстную кишку. Пропульсивные волны возникают с частотой 6- 7 в 1 мин. Они могут сочетаться и не сочетаться с перистальтическими.
Во время пищеварения сокращения продольной и циркулярной мускулатуры скоординированы и не отличаются друг от друга ни по форме, ни по частоте.
Регуляция моторной деятельности желудка. Регуляция двигательной активности желудка осуществляется центральными нервными, местными гуморальными механизмами. Нервная регуляция обеспечивается эффекгорными импульсами, поступающими к желудку по волокнам блуждающего (усиление сокращений) и чревных нервов (торможение сокращений). Афферентные импульсы возникают при раздражении рецепторов полости рта, пищевода, желудка, тонкой и толстой кишки. Адекватным раздражителем, вызывающим усиление двигательной активности мускулатуры желудка, является растяжение
410
его стенок. Это растяжение воспринимается отростками биполярных нервных клеток, расположенных в межмышечном и подслизистом нервных сплетениях.
Жидкости начинают переходить в кишку сразу после их поступления в желудок. Смешанная пища находится в желудке взрослого человека 3-10 часов.
Эвакуация пиши из желудка в двенадцатиперстную кишку обусловлена, в основном, сокращениями мускулатуры желудка — особенно сильными сокращениями его антрального отдела. Сокращения мускулатуры указанного отдела получили название пилорического "насоса". Градиент давления между полостями желудка и 12-ти перстной кишки при этом достигает 20-30 см вод. ст. Пилорический сфинктер (толстый циркуляторный слой мышц в области привратника) препятствует обратному забрасыванию химуса в желудок. На скорость опорожнения желудка влияют также величина давления в 12-ти перстной кишке, ее двигательная активность, величина рН содержимого желудка и двенадцатиперстной кишки.
В регуляции перехода пищи из желудка в кишечник первостепенное значение имеет раздражение механорецепторов желудка и двенадцатиперстной кишки. Раздражение первых ускоряет эвакуацию, вторых — замедляет ее. Замедление эвакуации наблюдается при введении в двенадцатиперстную кишку кислых растворов (с рН ниже 5,5), глюкозы, продуктов гидролиза жиров. Влияния этих веществ осуществляются рефлекторно, с участием "длинных" рефлекторных дуг, замыкающихся на различных уровнях центральной нервной системы, а также "коротких", нейроны которых замыкаются в экстра- и интрамуральных узлах.
Раздражение блуждающего нерва усиливает моторику желудка, увеличивает ритм и силу сокращений. При этом ускоряется эвакуация желудочного содержимого в 12-ти перстную кишку. Вместе с тем, волокна блуждающего нерва могут усиливать рецептивную релаксацию желудка и снижать моторику. Последнее происходит под влиянием продуктов гидролиза жира, действующих со стороны 12-ти перстной кишки.
Симпатические нервы снижают ритм и силу сокращений желудка, скорость распространения перистальтической волны.
Гастроинтестинальные гормоны также влияют на скорость эвакуации желудочного содержимого. Так, освобождение секретина и холецистокинин-панкреозимина под влиянием кислого содержимого желудка угнетает моторику желудка и скорость эвакуации из него пищи. Эти же гормоны усиливают панкреатическую секрецию, что вызывает повышение рН содержимого 12-ти перстной кишки, нейтрализацию хлористоводородной кислоты, т.е. создаются условия для ускорения эвакуации из желудка. Моторика усиливается так