Изотонический режим — сокращение, при котором мышца укорачивается без формирования напряжения. Такое сокращение возможно при пересечении или разрыве сухожилия или в эксперименте на изолированной (удаленной из организма) мышце. Изотоническое мышечное сокращение сокращение мышцы при неизменном напряжении, выражающееся в уменьшении её длины и увеличении поперечного сечения. В организме И. м. с. в чистом виде не наблюдается. К чисто И. м. с. приближается движение ненагруженной конечности; при постепенном увеличении груза до тех пор, когда он уже не может быть поднят, удаётся наблюдать все переходы от И. м. с. к изометрическому мышечному сокращению.
 14) Изометрическое мышечное сокращение
14) Изометрическое мышечное сокращение
Изометрический режим — сокращение, при котором напряжение мышцы возрастает, а длина практически не уменьшается. Такое сокращение наблюдается при попытке поднять непосильный груз.
Изометрическое мышечное сокращение сокращение мышцы, выражающееся в усилении её напряжения при неизменной длине (например, сокращение мышцы конечности, оба конца которой закреплены неподвижно). В организме к И. м. с. приближается напряжение, развиваемое мышцей при попытке поднять непосильный груз.
Механизм мышечного сокращения
Раздражение→ Возникновение потенциала действия→ Проведение его вдоль клеточной мембраны и в глубь волокна по поперечным трубочкам→ Освобождение Са2+ из боковых цистерн саркоплазматического ретикулума и диффузия его к миофибриллам→ Взаимодействие Са2+ с тропонином → Деформация тропомиозина → Взаимодействие поперечных мостиков с активными центрами актина → Скольжение актиновых нитей, приводящее к укорочению миофибриллы→ Активация кальциевого насоса→ Снижение концентрации свободны Са2+ в саркоплазме→ Отсоединение поперечных мостиков→ Расслабление миофибрилл

Саркомер
I- Строение мышечного волокна
II- Строение миофибриллы
A-Анизотропные темные диски
I-Изотропные светлые диски
H-зона, центральные части толстых миозиновых филаментов Z-линия, телофрагма
Описание и механизм:
Саркомер - функциональная единица сократительного аппарата миофибрилл мышечного волокна.
Сокращение- это процесс изменения длины или напряжения мышечного волокна при возбуждении. Саркомер состоит из тонких актиновых филаментов, расположенных в виде двух нитей бус, закрученных в двойную спираль. Между актиновыми нитями располагаются толстые миозиновые филаменты, имеющие две головки и длинный хвост. При генерации ПД в саркоплазматическом ретикулуме открываются кальциевые каналы, высвобождаются ионы кальция. Кальций соединяется с вспомогательным белком актиновых филаментов тропонином. Тропонин изменяет конфигурацию белка тропомиозина, который открывает ранее закрытые сайты связывания актина с миозином. Ионы кальция изменяют конфигурацию и миозиновых волокон. В результате их действие АТФ расщепляется до АДФ, в результате чего головка миозина присоединяется к актиновым бусинам, и тянет их в сторону М линии. Z линии сближаются, саркомер укорачивается (процесс сокращения). Как только возбуждение прекращается поперечные мостики распадаются за счет уменьшения концентрации кальция и настает расслабление, саркомер возвращается в исходную форму
 16) Структура нейрона
16) Структура нейрона
Нервная клетка (нейрон) – структурная и функциональная единица ЦНС, которая окружена клетками нейроглии.Он состоит из тела (сомы) и отходящих от негоотростков: аксона и дендритов. Скопление тел нейронов составляет серое вещество ЦНС, скопление отростков - белое вещество.
Тело нейрона покрыто плазматической мембраной (Мембрана тела покрыта синапсами, поэтому осуществляет восприятие и интеграцию импульсов, поступающих от других нейронов) и содержит в нейроплазме ядро и органоиды, в ней имеются и специфические образования – нейрофибриллы. Нейрофибриллы – тонкие опорные структуры, проходят в теле в различных направлениях,
продолжаются в отростки, располагаясь в них параллельно мембране. Они поддерживают определенную форму нейрона. Аксон – тонкий длинный отросток, покрытый миелиновой оболочкой. Функция аксона – проведение нервных импульсов от тела нейрона к другим нейронам или рабочим органам.
Дендриты – короткие, толстые ветвящиеся отростки, отходящие в большом количестве от тела нейрона (похожи на ветви дерева). Функция дендритов – восприятие раздражений или нервных импульсов от других нейронов и проведение их к телу нейрона.
Рецепторные (афферентные, чувствительные) нейроны проводят возбуждение (нервные импульсы) от рецепторов в ЦНС. Тела этих нейронов расположены в спинальных ганглиях, от тела отходит один отросток, который Т-образно делится на две ветви: аксон и дендрит. Дендрит (ложный аксон) – длинный отросток, покрыт миелиновой оболочкой, отходит от тела на периферию, разветвляется, подходя к рецепторам.
Эфферентные нейроны (командные по Павлову И.П.) проводят импульсы из ЦНС к органам, эту функцию выполняют длинные аксоны нейронов (длина может достигать 1,5 м.). Их тела располагаются в передних рогах (мотонейроны) и боковых рогах (вегетативные нейроны) спинного мозга.
Вставочные (контактные, интернейроны) нейроны – самая многочисленная группа, которые воспринимают нервные импульсы от афферентных нейронов и передают их на эфферентные нейроны. Различают возбуждающие и тормозящие вставочные нейроны.


17) I. Безмиелиновые волокна;
- возбуждение распространяется непрерывно.
II. Миелиновые волокна.
- возбуждение распространяется скачкообразно, т.е. сальтаторно.
Миелиновые волокна. Часть нервных волокон в ходе эмбриогенеза подвергается миелинизации: леммоциты (шванновские клетки) сначала прикасаются к аксону, а затем окутывают его. Мембрана леммоцита наматывается на аксон наподобие рулета, образуя многослойную спираль (миелиновую оболочку). Миелиновая оболочка не является непрерывной – по всей длине нервного волокна на равном расстоянии друг от друга в ней имеются небольшие перерывы (перехваты Ранвье). В области перехватов аксон лишен миелиновой оболочки.
Безмиелиновые волокна. Миелинизация других волокон заканчивается на ранних стадиях эмбрионального развития. В леммоцит погружается один или несколько аксонов; он полностью или частично окружает их, но не образует многослойной миелиновой оболочки
Механизм проведения возбуждения по миелиновым нервным волокнам
В миелинизированном нервном волокне участки мембраны, покрытые миелиновой оболочкой, являются невозбудимыми; возбуждение может возникать только в участках мембраны, расположенных в области перехватов Ранвье.
При развитии ПД в одном из перехватов Ранвье происходит реверсия заряда мембраны. Между электроотрицательными и электроположительными участками мембраны возникает электрический ток, который раздражает соседние участки мембраны. Однако в состояние возбуждения может перейти только участок мембраны в области следующего перехвата Ранвье. Таким образом, возбуждение распространяется по мембране скачкообразно (сальтаторно) от одного перехвата Ранвье к другому.
Механизм проведения возбуждения по безмиелиновым нервным волокнам
В состоянии покоя вся внутренняя поверхность мембраны нервного волокна несет отрицательный заряд, а наружная сторона мембраны – положительный. Электрический ток между внутренней и наружной стороной мембраны не протекает, так как липидная мембрана имеет высокое электрическое сопротивление.
Во время развития потенциала действия в возбужденном участке мембраны происходит реверсия заряда. На границе возбужденного и невозбужденного участка начинает протекать электрический ток. Электрический ток раздражает ближайший участок мембраны и приводит его в состояние возбуждения, в то время как ранее возбужденные участки возвращаются в состояние покоя. Таким образом, волна возбуждения охватывает все новые участки мембраны нервного волокна.
 18) Химический синапс. Синаптическая передача сигнала
18) Химический синапс. Синаптическая передача сигнала
1-Синаптический пузырек
2-Терминаль аксональная (пресинаптическая мембрана)
3-Синаптическая щель
4-Тело нервной клетки (постсинаптическая мембрана)
5-Нейромедиаторы
6-Рецепторы: холинорецепторы (М и N холинорецепторы),
адренорецепторы – α и β
Синапс – это структуры, обеспечивающие переход возбуждения с нервного волокна на иннервируемую им клетку.
Физиологические свойства химических синапсов:
1. Возбуждение передается при помощи медиаторов.
2. Обладают односторонним проведением возбуждения.
3. Быстрая утомляемость (истощение запасов медиатора).
4. Низкая лабильность 100-125 имп./сек.
5. Суммация возбуждения
6. Проторение пути
7. Синаптическая задержка (0,2-0,5 м/с).
8. Избирательная чувствительность к фармакологическим и биологическим веществам.
9. Чувствительны к изменениям температуры.
10. Существует следовая деполяризация.
Механизм передачи возбуждения в химических синапсах:
1. В возбуждающем синапсе (химический синапс):
импульс → нервное окончание в синаптические бляшки → деполяризация пресинаптической мембраны (вход Са++ и выход медиаторов) → медиаторы → синаптическая щель → постсинаптическая мембрана (взаимодействие с рецепторами) → генерация ВПСП → ПД.
2. В тормозных синапсах:
импульс → деполяризация пресинаптической мембраны → выделение тормозного медиатора → гиперполяризация постсинаптической мембраны (за счет К+) → ТПСП.
 19) Синапс
19) Синапс
1 – Строение синапса: 1 — нервное волокно; 2 — митохондрия; 3 — синаптический пузырек; 4 — предсинаптнческая мембрана; 5 — синаптическая щель; 6 — постсинаптическая мембрана; 7 — постсинаптические рецепторы
Синапс – это структуры, обеспечивающие переход возбуждения с нервного волокна на иннервируемую им клетку.
Рассмотрим строение синапса на примере аксо- соматического. Синапс состоит из трех частей: преси- наптического окончания, синаптической щели и пост- синаптической мембраны Пресинаптическое окончание (синаптическая бляшка) представляет собой расширенную часть тер- минали аксона. Синаптическая щель — это пространство между двумя контактирующими нейронами. Диаметр синаптической щели составляет 10 — 20 нм. Мембрана пресинаптического окончания, обращенная к синаптической щели, называется пресинаптической мембраной. Третья часть синапса — постсинаптичес- кая мембрана, которая расположена напротив пресинаптической мембраны. Пресинаптическое окончание заполнено пузырьками (везикулами) и митохондриями. В везикулах находятся биологически активные вещества — медиаторы. Медиаторы синтезируются в соме и по микротрубочкам транспортируются в пресинаптическое окончание. Наиболее часто в качестве медиатора выступают адреналин, норадреналин, ацетилхолин, серотонин, гамма-аминомасляная кислота (ГАМК), глицин и другие.
Синаптическая щель заполнена межклеточной жидкостью, в которой находятся ферменты, способствующие разрушению медиаторов. На одном постсинаптическом нейроне может находиться до 20000 синапсов, часть которых являются возбудительными, а часть — тормозными. Помимо химических синапсов, в которых при взаимодействии нейронов участвуют медиаторы, в нервной системе встречаются электрические синапсы. В электрических синапсах взаимодействие двух нейронов осуществляется посредством б
 химические синапсы. В некоторых межнейронных синапсах электрическая и химическая передача осуществляется одновременно — это смешанный тип синапсов. Влияние возбудительных и тормозных синапсов на возбудимость постсинаптического нейрона суммируется, и эффект зависит от места расположения синапса. Чем ближе синапсы расположены к аксональному холмику, тем они эффективнее. Напротив, чем дальше расположены синапсы от аксонального холмика (например, на окончании дендритов), тем они менее эффективны. Таким образом, синапсы, расположенные на соме и аксональном холмике, оказывают влияние на возбудимость нейрона быстро и эффективно, а влияние удаленных синапсов медленно и плавно. ПД нервного волокна (ПД — потенциал действия)
химические синапсы. В некоторых межнейронных синапсах электрическая и химическая передача осуществляется одновременно — это смешанный тип синапсов. Влияние возбудительных и тормозных синапсов на возбудимость постсинаптического нейрона суммируется, и эффект зависит от места расположения синапса. Чем ближе синапсы расположены к аксональному холмику, тем они эффективнее. Напротив, чем дальше расположены синапсы от аксонального холмика (например, на окончании дендритов), тем они менее эффективны. Таким образом, синапсы, расположенные на соме и аксональном холмике, оказывают влияние на возбудимость нейрона быстро и эффективно, а влияние удаленных синапсов медленно и плавно. ПД нервного волокна (ПД — потенциал действия)

Спиномозговые рефлексы
Спинной мозг организован по сегментарному принципу с сенсорными (чувствительными) корешками, входящими с дорзальной стороны каждого из 31 сегмента и моторными (двигательными) нервами, выходящими с вентральной стороны каждого сегмента.
Сегменты спинного мозга 8 шейных (C 1 - C 8) 12 грудных (Th 1 - Th 12) 5 поясничных (L 1 - L 5) 5 крестцовых (S 1 -S 5) 1-3 копчиковых (Co 1 - Co 2)
Участок спинного мозга, соответствующий двум парам корешков (два передних и два задних) называют сегментом СМ. Задние корешки - отростки межпозвоночных ганглиев оканчиваются у клеточных тел вставочных или двигательных нейронов. Передние корешки – двигательные – аксоны мотонейронов При сближении корешков образуется спинномозговой нерв.
Каждому определенному сегменту спинного мозга соответствует определенный участок кожи (дерматом), от которого берут начало нервные волокна, образующие задние корешки, и определенный раздел мускулатуры (миотом) тела, иннервируемый волокнами, составляющие передние корешки данного сегмента спинного мозга. Каждый сегмент спинного мозга иннервирует три поперечных отрезка - метамера тела (Шеррингтон).
Спинной мозг выполняет в основном две функции: Рефлекторную, Проводниковую.
Рефлекторная функция спинного мозга может быть сегментарной и межсегментарной. Рефлекторная сегментарная функция спинного мозга заключается в непосредственном регулирующем влиянии эфферентных нейронов спинного мозга на иннервируемые им эффекторы при раздражении рецепторов определенного дерматома.
Рефлексы дуга которых переключается в спинном мозге, называются спинальными. К простейшим спинальным рефлексам относятся сухожильные рефлексы, которые обеспечивают сокращение скелетных мышц при раздражении их проприорецепторов обусловленном быстрым
кратковременным растяжением мышцы (например, при ударе неврологическим молоточком по сухожилию). Сухожильные спинальные рефлексы являются клинически важными, т.к. каждый из них замыкается в определенных сегментах спинного мозга. Поэтому, по характеру рефлекторной реакции можно судить о функциональном состоянии соответствующих сегментов спинного мозга.
В зависимости от локализации рецепторов и нервного центра у человека различают:
Локтевой сгибательный рефлекс возникает при ударе по сухожилию двухглавой мышцы плеча (в области локтевой ямки) и проявляется в сгибании руки в локтевом суставе. Нервный центр этого рефлекса локализуется в 5-6 шейных сегментах спинного мозга.
Локтевой разгибательный рефлекс возникает при ударе по сухожилию трехглавой мышцы плеча (в области локтевой ямки) и проявляется в разгибании руки в локтевом суставе. Нервный центр этого рефлекса локализуется в 7-8 шейных сегментах спинного мозга.
Коленный рефлекс возникает при ударе по сухожилию четырехглавой мышцы бедра ниже коленной чашечки и проявляется в разгибании ноги в коленном суставе. Нервный центр этого рефлекса локализуется во 2-4 поясничных сегментах спинного мозга.
Ахиллов рефлекс возникает при ударе по пяточному сухожилию и проявляется в сгибании стопы в голеностопном суставе. Нервный центр этого рефлекса локализуется в 1-2 крестцовых сегментах спинного мозга.
Брюшной рефлексы: Верхний вызывается раздражением кожи под реберными дугами. Нервный центр этого рефлекса локализуется в 7-8 грудных сегментах спинного мозга. Средний вызывается штриховым раздражением кожи по направлению к пупку. Нервный центр этого рефлекса локализуется в 9-10 грудных сегментах спинного мозга. Нижний вызывают раздражением над пупартовой связкой. Нервный центр этого рефлекса локализуется в 11-12 грудных сегментах спинного мозга. Подошвенный рефлекс- кожный, выражающийся в сгибании стопы и пальцев при штриховом раздражении подошвы. Нервный центр этого рефлекса локализуется в 1-2 крестцовых сегментах спинного мозга.
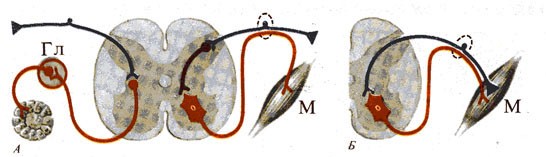
 21) Суммация возбуждений
21) Суммация возбуждений
Различают два вида суммации:
- пространственная (а, б)
- последовательная или временная (а, б, в)
В ответ на одиночную афферентную волну, идущую от рецепторов к нейронам, в пресинаптической части синапса освобождается небольшое количество медиатора. При этом в постсинаптической мембране нейрона обычно возникает ВПСП-небольшая местная деполяризация. Для того чтобы общая по всей мембране нейрона величина ВПСП достигала порога возникновения потенциала действия, требуется суммация на мембране клетки многих подпороговых ВПСП. Лишь в результате такой суммации возбуждения возникает ответ нейрона.
Пространственная суммация наблюдается в случае одновременного поступления нескольких импульсов в один и тот же нейрон по разным пресинаптическим волокнам. Одномоментное возбуждение синапсов в различных участках мембраны нейрона повышает амплитуду суммарного ВПСП до пороговой величины. В результате возникает ответный импульс нейрона и осуществляется рефлекторная реакция.
Временная суммация происходит при активации одного и того же афферентного пути серией последовательных раздражений. Если интервалы между поступающими импульсами достаточно коротки и ВПСП нейрона от предыдущих раздражений не успевают затухать, то последующие ВПСП накладываются друг на друга, пока деполяризация мембраны нейрона не достигает критического уровня для возникновения потенциала действия. Таким способом даже слабые раздражения через некоторое время могут вызвать ответные реакции организма.
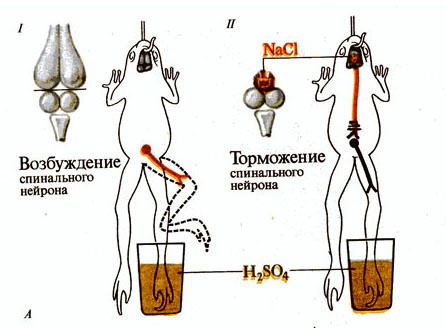 22) Торможение в ЦНС. Опыт Сеченова
22) Торможение в ЦНС. Опыт Сеченова
I — двигательный рефлекс на болевой раздражитель
II — распространение нервных импульсов от тормозных нейронов ствола мозга к спинному мозгу при наложении кристалла NaCI на
область зрительных чертогов и отсутствие двигательного рефлекса на болевой раздражитель.
Интегративная деятельность в ЦНС осуществляется при участии возбуждающих и тормозных процессов.
Явление торможения в 1863 году открыл акад. И.М. Сеченов (опыт рисунок стр. 239). Ч. Шеррингтон, Н. Е. Введенский, А.А. Ухтомский, И.П. Павлов показали, что торможение имеет место в работе всех отделов мозга.
Торможение – активный процесс, проявляющийся внешне в подавлении или ослаблении процесса возбуждения.
Торможение в ЦНС (И.М. Сеченов) - активный самостоятельный нервный процесс, проявляющийся внешне в подавлении или ослаблении процесса возбуждения и характеризующийся определенной интенсивностью и длительностью.
Классический опыт Сеченова заключается в следующем: у лягушки с перерезанным головным мозгом на уровне зрительных бугров определяли время сгибательного рефлекса при раздражении лапки серной кислотой. После этого на зрительные бугры накладывали кристаллик поваренной соли и снова определяли время рефлекса. Оно постепенно увеличивалось, вплоть до полного исчезновения реакции. После снятия кристаллика соли и промывания мозга физиологическим раствором время рефлекса постепенно восстанавливалось. Это позволило говорить о том, что торможение — активный процесс, возникающий при раздражении определенных отделов ЦНС.