(двух нейронная)
Рефлекторная дуга – последовательно соединенная цепочка нервных клеток, которая обеспечивает осуществление реакции, ответа на раздражение. Она состоит из двух половин: 1) чувствующей, или приводящей и 2) двигательной, или отводящей.
1. Вегетативная нервная система обеспечивает регуляцию всех функций внутренних органов, сосудов, потовых желез, а также трофику скелетных мышц.
ВНС подразделяется на симпатическую (от шеи до поясничного отдела) и парасимпатическую (от продолговатого мозга до крестцового отдела) (СНС, ПНС).
Центры ВНС имеют очаговый выход: - центры парасимпатического отдела ВНС располагаются в продолговатом, среднем мозге и крестцовом отделе спинного мозга. - центры симпатического отдела ВНС находятся только в спинном мозге (грудной и 1-4 поясничные сегменты)
Эфферентное звено ВНС двух нейронный. Тело первого нейрона в ЦНС, второй в ганглиях. Ганглии СНС в зависимости от локализации делятся:
- паравертебальные (симп. цепочк.)
- превертебральные. Ганглии ПСН располагаются внутриорганов(интрамуральные).
Волокна делятся преганглионарные и постганглионарные. Преганглионарные волокна миелиновые, постганглионарные безмиелиновые, типа В,С.
Симпатическая нервная система: преганглионарные волокна короткие, а постганглионарные длинные.
Парасимпатическая нервная система: преганглионарные волокна длинные, а постганглионарные короткие.
Длительная синаптическая задержка – 1,5 – 30 мс.
ПД длительный, низкая возбудимость, большой рефрактерный период, большая хронаксия.
Длительный следовой потенциал (300-400 мс.) поэтому вегетативные рефлексы протекают интерно.
Основным способом передачи возбуждения в синапсах является химический способ. Синапс выделяет только один медиатор (принцип Дейла), действие медиатора зависит от природы рецептора постсинаптической мембраны.
Медиаторы: Ацетилхолин (превертебральные ганглии), Норадреналин (потовые железы, кожа, сосуды, скелетная мускулатура).
Путь: импульс-рецептор-афферентный путь-чувствительный нейрон-задние корешки спинного мозга-эфферентный нейрон (боковые корешки)-передние корешки-ганглий-эффектор.
2. Сомати́ческая не́рвная систе́ма (от греч. soma — тело) — часть нервной системы человека, представляющая собой совокупность афферентных (чувствительных) и эфферентных (двигательных) нервных волокон, иннервирующих мышцы (у позвоночных — скелетные), кожу, суставы. Эта система состоит из нервов, прикрепленных к коже, органам чувств и всем мышцам скелета.
Соматическая рефлекторная дуга является трех нейронной.
Нейроны соматической нервной системы тянутся от центральной нервной системы прямо к мускулам и рецепторам. Тело нейрона находится в центральной нервной системе, а аксоны тянутся дальше, пока не достигают кожи, органов чувств или мышц.
Путь: импульс-рецептор (экстрацептивный рецептор)-афферентный путь-чувствительный нейрон-задние корешки спинного мозга вставочный нейрон- двигательный нейрон передние корешки спинного мозга-эфферентный путь-эффектор.
3. Сухожильный рефлекс - рефлекторное сокращение скелетной мышцы или их групп в ответ на раздражение проприорецепторов сухожилия одной из этих мышц.
Сухожильная рефлекторная дуга содержит 2 нейрона.
Путь: импульс-рецептор (проприорецептор) –афферентный путь-чувствительный нейрон-задние корешки-мотонейрон-передние корешки-эффферентный путь-эффектор.
 24) Торможение по типу пессимума Н.Е. Введенского
24) Торможение по типу пессимума Н.Е. Введенского
Пессимальное торможение
Представляет собой вид торможения центральных нейронов. Наступает при высокой частоте раздражения. Не требует специальных структур. В основе лежит механизм инактивации Na -каналов при длительной деполяризации и изменение свойств мембраны аналогично катодической депрессии. (Пример - лягушка, перевернутая на спину – мощная афферентация от вестибулярных рецепторов – явление оцепенения, гипноза). Не требует специальных структур. Торможение обусловлено выраженной следовой гиперполяризацией постсинаптической мембраны в аксональном холмике после длительного возбуждения.
Пессимальное торможение
Для ЦНС свойственно пессимальное торможение, близкое по своему происхождению пессимуму Введенского в нервно-мышечных соединениях. Пессимальное торможение развивается в синапсах при разражениях высокой частоты и зависит от низкой лабильности синапсов. Данное явление связано с появлением при частых импульсах стойкой деполяризации постсинаптической мембраны, что нарушает распространение возбуждения и рефлекс тормозится. К пессимальному торможению особенно склонны вставочные нейроны спинного мозга и клетки ретикулярной формации. Пессимальное торможение предохраняет нервную клетку от перевозбуждения.
В нервной клетке может развиваться торможение после прекращения ее возбуждения – так называемое торможение вслед за возбуждением, возникающее в том случае, если после окончания возбуждения в клетке развивается сильная следовая гиперполяризация мембраны. Если в этот момент к клетке приходит новый импульс, то возникший посгсипаптический потенциал оказывается недостаточным для критической деполяризации мембраны.
 25) Принцип реципрокности
25) Принцип реципрокности
Принцип реципрокности – согласованная работа нервных центров, особенно у противоположных рефлексов (сгибание, разгибание и т. д.).
Принцип реципрокности (сопряженности) в работе нервных центров. Суть его заключается в том, что при возбуждении одних нервных центров деятельность других может затормаживаться. Принцип реципрокности был показан по отношению к нервным центрам мышц-антагонистов - сгибателей и разгибателей конечностей. Только при такой взаимосочетанной (реципрокной) иннервации возможен акт ходьбы. Наиболее отчетливо он проявляется у животных с удаленным головным мозгом и сохраненным спинным (спинальное животное), но может происходить сопряженное, реципрокное торможение и других рефлексов. Под влиянием головного мозга реципрокные отношения могут изменяться. Человек или животное в случае необходимости может сгибать обе конечности, совершать прыжки и т. д. Реципрокные взаимоотношения центров головного мозга определяют возможность человека овладеть сложными трудовыми процессами и не менее сложными специальными движениями, совершающимися при плавании, акробатических упражнениях и пр. Принцип отражает характер отношений между центрами, ответственными за осуществление противоположных функций (вдоха и выдоха, сгибание и разгибание конечностей), и заключается в том, что нейроны одного центра, возбуждаясь, тормозят нейроны другого и наоборот.
 26) Дивергенция
26) Дивергенция
– способность НЦ устанавливать многочисленные синаптические связи с различными клетками. Благодаря этому НК могут участвовать в нескольких рефлекторных актах. Таким образом между НЦ отсутствует жесткая стабильность и координационные отношения могут меняться и в результате возникает адекватная рефлекторная реакция.
Дивергенция - это способность нейрона устанавливать многочисленные связи с другими нейронами. Вследствие этого одна и та же клетка может участвовать в различных нервных процессах и реакциях, контролировать большое число других нейронов, то есть каждый нейрон может обеспечить распространение импульсов - иррадиацию возбуждения. Процессы дивергенции более типичны для афферентных отделов ЦНС.
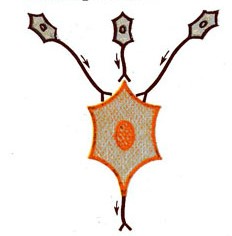 27) Конвергенция
27) Конвергенция
Конвергенция – схождения импульсов разной модальности на одних и тех же нервных центрах.
На каждом из нейронов ЦНС конвергирует (сходятся) различные афферентные волокна. Таких афферентных входов для большинства нейронов много десятков и даже тысяч. Так, на мотонейронах заканчиваются в среднем 6000 коллатералей аксонов, которые поступают от периферических рецепторов и различных структур мозга, образуя возбуждающие и тормозные синапсы. Это такое универсальное явление, можно говорить о принципе конвергенции в нейронах и их связях. Благодаря этому явлению в один и тот же нейрон одновременно поступают многочисленные и разнообразные потоки возбуждений, которые затем подлежат сложной обработке и перекодируются и формируются в единое возбуждение - аксону, что идет к следующему звену нервной сетки. Конвергенция возбуждения на нейроне является универсальным фактором его интегративной деятельности. Различают: мультисенсорную, мультибиологическую и сенсорно-биологическую формы конвергенции. В первом случае на нейрон поступают сигналы различной сенсорной модальности (зрительные, слуховые, болевые и др.), во втором - потоки возбуждений различной биологической модальности (пищевые, половые и др.), в третьем - сигнализация (зрительная, пищевая) и другие.
 28) Принцип общего конечного пути (воронки), установил Ч. Шеррингтон
28) Принцип общего конечного пути (воронки), установил Ч. Шеррингтон
Английский физиолог Ч. Шеррингтон установил, что одна и та же двигательная реакция может быть получена при раздражении различных афферентов. Это обусловлено тем, что отдельные мотонейроны являются общим конечным путем многочисленных сенсорных каналов. Принцип общего конечного пути («воронка»Шеррингтона) заключается в конкуренции множества афферентов и интернейронов за «обладание» одним общим конечным мотонейроном. Возбуждения различной сенсорной модальности и биологического качества конвергируют к одному мотонейрону. Из всего поступающего потока мотонейрон отбирает приоритетную (наиболее значимую) информацию и синтезирует эфферентную команду для исполнителей. При этом отбор приоритетной информации в ходе «борьбы» афферентов за «обладание» общим конечным мотонейроном обусловлен окклюзией возбуждений в нервных центрах. Принцип конвергенции возбуждения (или принцип общего конечного пути, воронка Шеррингтона). Конвергенция нервных импульсов (от лат. convergo, convergere - сближать, сходиться) означает схождение к одному нейрону двух или нескольких различных возбуждений одновременно. Это явление было открыто Ч. Шеррингтоном. Он показал, что одно и то же движение, например рефлекторное сгибание конечности в коленном суставе, можно вызвать путем раздражения различных рефлексогенных зон. В связи с этим им было введено понятие "общего конечного пути", или "принципа воронки", согласно которому потоки импульсов от различных нейронов могут сходиться на одном и том же нейроне (в данном случае - на альфа-мотонейронах спинного мозга). В частности, Ч. Шеррингтон обнаружил схождение к одним и тем же промежуточным или эфферентным нейронам различных афферентов от разных участков общего рецептивного поля (в спинном и продолговатом мозге) или даже от разных рецептивных полей (в высших отделах головного мозга). В настоящее время показано, что конвергенция возбуждения, так же как и дивергенция возбуждения, - очень распространенное явление в ЦНС. Основой для конвергенции (как и для иррадиации) является определенная морфологическая и функциональная структура различных отделов мозга. Очевидно, что часть конвергентных путей является врожденными, а другая часть (главным образом в коре большого мозга) - приобретенная в результате обучения в процессе онтогенеза. Формирование новых конвергентных отношений для нейронов коры большого мозга в процессе онтогенеза во многом связаны с формированием в коре доминантных очагов возбуждения, которые способны "притягивать" к себе возбуждение от других нейронов.
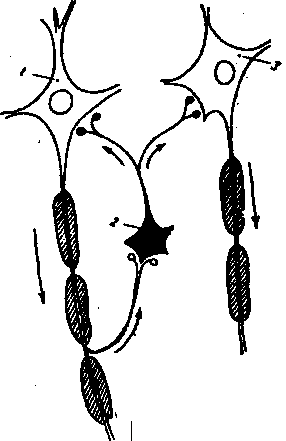 29) Схематическое изображение связен между мотонейронами и клетками Рэншоу.
29) Схематическое изображение связен между мотонейронами и клетками Рэншоу.
Коллатерали аксона мотонейрона (1) контактируют с телом клетки Рэншоу (2), короткий аксон которой, разветвляясь, образует тормозные синапсы на мотонейроне 1 и мотонейроне 3.
Иррадиация возбуждения
Импульсы, поступающие в центральную нервную систему при сильном длительном раздражении, вызывают возбуждение не только нейронов данного рефлекторного центра, но и нервных центров. Это распространение возбуждения в центральной нервной стеме получило название иррадиации.
Для иллюстрации приведем результаты следующего опыта. Если у кошки после перерезки мозгового ствола, отделяющей вышележащие отделы головного мозга (например, большие полушарии) от нижележащих, нанести слабое раздражение на подошву задней лапы, то наблюдается сгибание только этой лапы в голеностопном суставе. Усиление раздражения приводит к сгибанию, кроме того, ещё и в коленном суставе, а еще большее усиление - к сгибанию и в тазобедренном суставе. Еще более сильное раздражение вызывает, кроме перечисленных сгибательных движений данной конечности, разгибание задней конечности противоположной стороны. При продолжающемся усилении раздражения происходит разгибание передней конечности одноимённой стороны и, наконец, сгибание симметричной передней конечности.
Морфологические и электрофизиологические исследования показали, что возможность иррадиации возбуждений в центральной нервной системе обусловлена многочисленными ветвлениями отростков (аксонов и дендритов) нервных клеток и цепями вставочных нейронов, объединяющих друг с другом различные нервные центры. Особую роль в механизме иррадиации возбуждения играет ретикулярная, формация.
Иррадиации возбуждения препятствуют многочисленные тормозные нейроны и синапсы, входящие в состав различных рефлекторных центров.
Значение торможения в ограничении иррадиации возбуждения хорошо иллюстрирует опыт с введением животному стрихнина, блокирующего тормозные синапсы в центральной нервной системе и тем самым устраняющего постсинаптическое торможение Если под кожу лягушке ввести 0,1% раствор стрихнина, то уже через несколько минут достаточно дотронуться к лапке лягушки или даже постучать по столу, на котором она сидит, чтобы возникло сильнейшее общее возбуждение центральной нервной системы, которое сопровождается судорогами всех скелетных мышц. Опыт показывает, что выключение тормозных синапсов имеет следствием широкую иррадиацию возбуждения в цептральной нервной системе.
Аналогичные явления наблюдаются и при воздействии на организм столбнячного токсина, также выключающего тормозные синапсы.
В механизме ограничения иррадиации возбуждения имеют большое значение нервные клетки, открытые Б. Реншоу в спинном мозгу. Этот исследователь показал, что аксоны мотонейронов перед выходом из спинного мозга часто дают одну или несколько коллатералей. Последние оканчиваются
на клетках Реншоу, аксоны которых образуют тормозные синапсы на мотонейронах данного сегмента спинного мозга (рис. 183).
Благодаря этому возбуждение, возникающее в мотонейроне, по прямому пути распространяется на периферию к скелетной мышце, а по коллатерали — активирует тормозную клетку, которая подавляет возбуждение мотонейрона.
Чем интенсивнее возбуждение мотонейрона и, следовательно, чем больше частота импульсов, посылаемых им на периферию, тем сильнее возбуждается клетка Реншоу, которая тормозит активность мотонейрона. Таким образом, существует механизм, автоматически охраняющий нервные клетки от их чрезмерного возбуждения.
Торможение, осуществляемое при участии клеток Реншоу, получило название возвратного торможения. Как показали недавние исследования Дж. Экклса и др., оно имеет значение в деятельности всех отделов центральной нервной системы.
 30) Пресинаптическое торможение
30) Пресинаптическое торможение
Расположение тормозных синапсов на пресинаптических разветвлениях аксонов (схема).
Н — нейрон, возбуждаемый афферентными импульсами, приходящими по волокну 1;
Т — нейрон, образующий тормозные синапсы на пресинаптических разветвлениях волокна 1;
2 —афферентные волокна, вызывающие активность тормозного нейрона Т
Пресинаптическое торможение, как это следует уже из самого названия локализуется в пресинаптических элементах, а именно в тончайших разветвлениях аксонов перед их переходом в нервное окончание.
На этих разветвлениях — пресинаптических миналях — располагаются окончания других нервных клеток, образующие здесь особые тормозные синапсы (рис. 181).
Медиаторы, выделяющиеся в этих тормозных синапсах, деполяризуют в мембрану терминалей и приводят их в стояние, подобное катодической депрессии Вериго. Это обусловливаетчастичную или полную блокаду проведения нервных импульсов к нервным окончаниям.
Пресинаптическое торможение первоначально выявлено в спинном мозге в опыте с регистрацией активности мотонейронов моносинаптической рефлекторной дуги при раздражении антагонистических мышечных нервов. Так, известно, что раздражение первичных афферентов мышечных веретен сопровождается возбуждением гомонимных а-мотонейронов (а-мотонейро-нов этой же мышцы). Однако опережающее раздражение афферентов сухожильных рецепторов мышц-антагонистов предотвращает возбуждение активируемых а-мотонейронов. Интересным в данном случае было то, что мембранный потенциал и возбудимость исследуемых а-мотопейронов не изменя-лись либо регистрировался низкоамплитудный ВПСП, недостаточный для возникновения ПД. В опыте исследовались мотонейроны в составе моносинаптической рефлекторной дуги, поэтому было очевидно: они не возбуждаются вследствие процессов, происходящих в пресинаптическом окончании, что определяет название этого вида торможения.
 31) Центральное облегчение
31) Центральное облегчение
А- При раздражении А возбуждаются 2 нейрона (1, 2)
В- При раздражении В возбуждаются 2 нейрона (5, 6)
Темный овал - Клетки периферической каймы
Светлый - Клетки центральной части нейронного пула
?-При раздражении А+В возбуждаются все 6 нейронов
Центральное облегчение - объясняется особенностями строения нервного центра. Каждое афферентное волокно входя в нервный центр иннервирует определенное количество нервных клеток. Эти нейроны - нейронный пул. В каждом нервном центре много пулов. В каждом нейронном пуле - 2 зоны: центральная (здесь афферентное волокно над каждым нейроном образует достаточное для возбуждения количество синапсов), периферическая или краевая кайма (здесь количество синапсов недостаточно для возбуждения). При раздражении возбуждаются нейроны центральной зоны.
Центральное облегчение: при одновременном раздражении 2-х афферентных нейронов ответная реакция может быть больше арифметической суммы раздражения каждого из них, т. к. импульсы от них отходят к одним и тем же нейронам периферической зоны.
 32) Центральная окклюзия
32) Центральная окклюзия
А- При раздражении А возбуждаются 4 нейрона (1, 2, 3, 4)
В- При раздражении В возбуждаются 4 нейрона (3, 4, 5, 6)
Круг - клетки центральной части нейронного пула
?- При раздражении А+В возбуждаются все 6 нейронов
Окклюзия - уменьшение эффекта одновременного возбуждения двух сильных афферентных входов, по сравнению с суммой эффектов при раздельном их раздражении.
Причина окклюзии заключается в том, что эти афферентные входы в результате конвергенции адресуются к совместных '' пороговых " мотонейронов и каждый может создать в них такое же надпороговая возбуждения, как и оба входа вместе.
 33) Последействие
33) Последействие
Схема кольцевых связей в нервном центре
(по Лоренто де Но):
1 — афферентный путь;
2-промежуточные нейроны;
3 — эфферентный нейрон;
4 — эфферентный путь;
5 — возвратная ветвь аксона Последействие - способность сохранять возбуждение после окончания действия раздражителя, т.е. афферентных импульсов нет, а эфферентные продолжают действовать еще некоторое время.
Последействие объясняется наличием следовой деполяризации. Если следовая деполяризация длительна, то на ее фоне в течение нескольких миллисекунд могут возникать потенциалы действия (ритмическая активность нейрона), вследствие чего сохраняется ответная реакция. Но это дает сравнительно короткий эффект последействия. Более длительное последействие связано с наличием кольцевых связей между нейронами. В них возбуждение как бы само себя поддерживает, возвращаясь по коллатералям к первоначально возбужденному нейрону
 34) Схема симпатической нервной системы
34) Схема симпатической нервной системы
1- Спинной мозг
2- Симпатический ствол
3- Узловой нервный ганглий
4- Грудные спинномозговые сегменты
5- Поясничные спинномозговые сегменты
6- Преганглионарные нервные волокна
7- Постганглионарные нервные волокна (штрихованные - первая группа; сплошные- вторая группа)
8- Место разветвления обособленного стволика второй группы постганглионарных волокон.
9- Волокна нервного узла, идущие к исполнительному органу
На данном рисунке изображена простая схема строения СИМПАТИЧЕСКОЙ нервной системы, являющейся частью ВНС. Красным цветом изображен спинной мозг, а закрашенные красным ячейки- сегменты участвующие в образовании симпатического нервного ствола и содержащие симпатические ядра, а именно CVII-CVIII;ThI-ThXII; LI-LIII.Это центральный отдел СНС.
Зеленым изображен сам симпатический ствол, который соединен со спинным мозгом посредством ПРЕГАНГЛИОНАРНЫХ волокон, которые в отличие от Парасимпатической НС короткие. Далее нервный импульс через эфферентные нейроны симпатического ствола, либо проводиться к другим эфферентным нейронам в пределах симпатического ствола, либо продолжает движение в ПОСТГАНГЛИОНАРНЫХ ВОЛОКНАХ. Данные волокна образуют две группы: волокна которые проводят импульс непосредственно к рабочему органу или обратно к спинномозговому нерву и волокна, проводящие импульс к периферическим эфферентным нейронам, входящих в состав нервных узлов. Через 3 эфферентный нейрон импульс движется к исполнительному органу.
 35) Схема парасимпатической нервной системы
35) Схема парасимпатической нервной системы
1- средний мозг
2- продолговатый мозг
3- крестцовый отдел спинного мозга
4- эфферентный ганглий
5- преганглионарное волокно
6- постганглионарное волокно
На данном рисунке описана простая схема строения ПАРАСИМПАТИЧЕСКОЙ НС. Красным нарисован ствол головного мозга и спинной мозг. Синим заштрихованы: 1- средний мозг, от которого отходят парасимпатические волокна глазодвигательного нерва III, регулирующие рефлек
и расширение зрачка; 2- Продолговатый мозг от которого отходят парасимпатические волокна ЛИЦЕВОГО VII ЯЗЫКОГЛОТОЧНОГО IX БЛУЖДАЮЩЕГО X нервов; 3- крестцовый отдел спинного мозга от которого берет начало тазовый нерв.
На схеме видно, что парасимпатические волокна не образуют предпозвоночного ствола, а ПРЕГАНГЛИОНАРНЫЕ волокна длиннее, чем у волокон СИМПАТИЧЕСКОЙ НС, а эфферентный ганглий расположен ближе к исполнительному органу
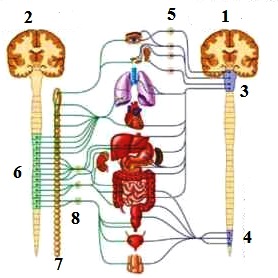 36) Строение вегетативнй нервной системы
36) Строение вегетативнй нервной системы
1- Парасимпатическая
2- Симпатическая нервная система
3- Ствол головного мозга
4- крестцовый отдел спинного мозга
5- эфферентный ганглий
6- отделы спинного мозга участвующие в симпатической иннервации
7- симпатический ствол
8- преганглионарные и постганглионарные нервные волокна.
На рисунке изображен общий упрощенный план строения ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ.
Слева изображена схема строения СИМПАТИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ, а справа ПАРАСИМПАТИЧЕСКАЯ НЕРВНАЯ СИСТЕМА. В каждой из них выделяют ЦЕНТРАЛЬНЫЙ и ПЕРИФЕРИЧЕСКИЙ отделы. Центральный отдел СНС представлен вегетативными ядрами, расположенными в концевых шейных, всех грудных и начальных сегментах спинного мозга от которых отходят преганглионарные нервные волокна, которые затем входят в симпатический ствол. От симпатического ствола начинается периферический отдел, который представлен постганглионарными волокнами, которые отходят от симпатического ствола и движутся, либо непосредственно к исполнительным органам, либо через ганглий нервных сплетений и узлов к исполнительны органам.
ПАРАСИМАТИЧЕСКАЯ НС представлена нервными волокнам, отходящими от далеко расположенных относительно друг друга отделов ствола головного мозга и спинного мозга, а именно Средний, продолговатый мозг и крестцовый отдел спинного мозга от которых отходят парасимпатические волокна. Это центральный отдел. Преганглионарные волокна в отличие от СНС длиннее, а эфферентный ганглий расположен ближе к исполнительному органу.

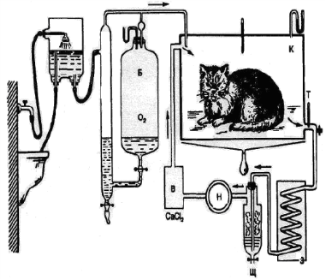
Прямая калориметрия
Биокалориметр Этуотера – Бенедикта
На данной схеме изображен принцип действия ПРЯМОЙ КАЛОРИМЕТРИИ. ПРЯМАЯ КАЛОРИМЕТРИЯ - метод исследования обменных процессов человека, основанный на измерении выделенного телом человека количества тепла за определенный промежуток времени. Данная процедура проводится в специальной камере. Данная камера оборудована системой трубок по которым течет вода из резервуара 3. Вода поглощая тепло изменяет температуру, которая фиксируется термометрами 1 и 2. Система вентиляции камеры 5 пропускает воздух через емкости 6,7,8 в которых находятся: вещество поглощающее воду(серная кислота), вещество поглощающее углекислый газ (натриевая известь). Кислород в камеру подается
из баллона 10, а затраченное его число фиксируется газовыми часами 11. Через окошко 4 испытуемому доставляется пища. Под номером 12 обозначен регулятор атмосферного давления в камере. Данный метод достаточно точен, но сложен в выполнении и не позволяет измерить энергозатраты при выполнении умственной и физической работы.
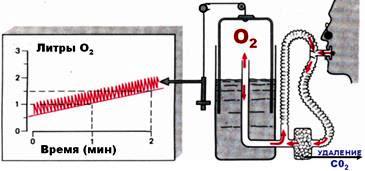
Непрямая калориметрия
Данные рисунки демонстрируют метод НЕПРЯМОЙ КАЛОРИМЕТРИИ. Данный метод измерения обменных процессов основан на определении количества кислорода, затраченного на окисление органических веществ по отношению к поглощенному кислороду, а также количеству углекислогогаза, выделенного в результате окисления.
Непрямая калориметрия методом КРОГА.
Испытуемый подключается к спирометаболографу. Через маску он вдыхает ЧИСТЫЙ КИСЛОРОД из резервуара. Часть кислорода поглощается и преобразуется в углекислый газ, а неиспользованный кислород остается. После выдоха неиспользованный кислород возвращается в резервуар, а углекислый газ удаляется химическим методом. Прибор фиксирует объем поглощенного кислорода во времени и по уже известным значениям КАЛОРИМЕТРИЧЕСКОГО КОЭФФИЦИЕНТА КИСЛОРОДА для определенных питательных веществ, производится вычисление основного обмена.

Метод Дугласа-Холдена
Метод Дугласа- Холдена. Испытуемый надевает маску, которая устроена таким образом что он может вдыхать атмосферный воздух, а выдыхаемый воздух попадает в специальный мешок закрепленный за спиной. Мешок оборудован газовыми часами которые фиксируют количество поступившего выдохнутого воздуха. В течение определенного времени испытуемый выполняет определенную работу. По истечению времени испытания, выдохнутый воздух подвергается физико-химическому исследованию для определения точного количества углекислого газа и неиспользованного кислорода. По отношению выделенного углекислого газа к количеству поглощенного кислорода вычисляется дыхательный коэффициент.