Возникновение возбуждения и генерация потенциала действия (ПД) происходят в том случае, если раздражение, нанесенное на возбудимую клетку, снижает ее мембранный потенциал (МП) до определенного уровня — критического уровня деполяризации. Такая реакция на сдвиг МП (на деполяризацию) возможна благодаря тому, что натриевая проводимость мембраны (gNa) является функцией МП. При нормальном потенциале покоя величина (gNa) очень мала, снижение МП ведет к ее росту, в результате чего большее количество Na+ проникает в клетку и ускоряется перенос положительных зарядов внутрь клетки, что способствует еще большему снижению уровня потенциала покоя. Это, в свою очередь, ведет к увеличению натриевой проводимости и т. д. Быстро протекающее колебание МП (1-ьЗмс) — пик, или спайк достигает вели; чины 100-:-120 мВ. Теоретически при возбуждении МП должен достигнуть уровня натриевого равновесного потенциала, уравновешивающего натриевый концентрационный градиент VNa = 58 Ig(Na1)H/(Na+)e. Однако практически этот уровень никогда.не достигается вследствие развивающейся натриевой инактивации и действия восстанавливающего фактора — возрастания калиевой проводимости (gK) мембраны при деполяризации. Рост ^к)мембраны приводит к достаточно быстрому возвращению смещенного МП к исходному уровню потенциала покоя (рис. 7.1). Для миелинизированных нервных волокон характерно замедление возврата МП в нескольких милливольтах от уровня покоя. Это отрицательный следовой потенциал. Его развитие связывают с некоторым накоплением ионов К+, вышедших из аксонов, около мембраны перехвата Ранвье благодаря слоям окружающих клеток. У других объектов (например, гигантский аксон кальмара) регистрируют положительный следовой потенциал — следовую гиперполяризацию. Ее связывают с временно увеличенной проницаемостью для Ю. '
Нервные и мышечные волокна обладают кабельными свойствами: протоплазма окружена поверхностной мембраной, изолирующей этот цилиндрический проводник от электролитов межклеточной жидкости (рис.8). На рис. 8 представлена общая схема строения нервной клетки. Из-за утечки тока через емкость и сопротивление мембраны подпорого-вые стимулы распространяются всего на 1-н2 мм вдоль волокна. Благодаря регенеративному характеру деполяризации мембраны пороговый сдвиг МП подвергается 4- или 5-кратному усилению. Это компенсирует недостатки кабельной структуры нервного или мышечного волокна и
ПД распространяется по всей длине волокна без уменьшения амплитуды. Скорость его распространения, или скорость проведения возбуждения, зависит от внутреннего и внешнего сопротивления клетки, сопротивления и емкости ее мембраны, от амплитуды ПД.
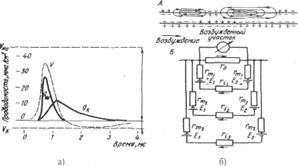
Рис. 7.
Рис. 7а. Кривые изменения натриевой, калиевой проводимости и возникающего
при этом потенциала. Рис. 76. Местные токи возбужденного аксона (А)
и схема регистрации ПД при внеклеточном отведении (Б)
При внеклеточном отведении от нервного ствола,(группы аксонов) регистрируют не изменение потенциала на мембране, а падение напряжения на внеклеточном межэлектродном участке, вызванное протеканием локальных токов вдоль наружной поверхности множества аксонов от покоящихся участков к активному (рис.7.2). Именно с этим связан рост амплитуды регистрируемого внеклеточно ПД при увеличении числа возбужденных аксонов.
Возбуждение нервных и мышечных клеток сопровождается рефрак-терностью (рис. 9).

|

|

|
Рис. 8. Схема строения нервной клетки

Рис.9. Кривые (1-4), демонстрирующие восстановление ПД по мере окончания рефрактерного периода. Кривая (5) — реконструкция этого процесса
Генерация второго ПД во время нисходящей фазы предыдущего спайка невозможна из-за развития натриевой инактивации (нет повышения (gNa)при деполяризации) и увеличения (§ка). которая становится значительно выше натриевой. Поэтому весь период спайка ПД нерв невозбудим — абсолютный рефрактерный период. После окончания спайка натриевая инактивация исчезает не мгновенно, а в течение определенного временного интервала. Повышенная калиевая проводимость тоже не сразу возвращается к уровню покоя. В связи с этим в течение некоторого периода после спайка порог раздражения увеличен (понижение возбудимости) и амплитуда генерируемых в этот период ПД ниже нормальной. Это относительный рефрактерный период.
Для измерения длительности рефракторного периода используют сдвоенные раздражающие стимулы, временной интервал между которыми можно изменять. 1-й стимул St называют кондиционирующим, В ответ на него возникает нормальный ПД, который служит эталоном для сравнения, 2-й раздражающий стимул $2 называется тестирующим, ибо он помогает определить возбудимость нерва через различные временные интервалы после первого ПД. Об окончании абсолютной рефрактерности судят по появлению ПД на тестирующий стимул при по-
степенном увеличении интервала между 1-м и 2-м стимулами от нуля. Об окончании относительной рефрактерности судят по восстановлению либо амплитуды эталонного ПД, либо прежнего порога раздражения 4при дальнейшем увеличении интервала между 1-м и 2-м стимулами. Кроме того, при внеклеточном отведении регистрирующие электроды расположены на нерве достаточно близко друг к другу по сравнению с длиной волны возбуждения. Последняя равна произведению скорости проведения на длительность возбуждения в данной точке нерва. Так, если принять для седалищного нерва лягушки скорость проведения порядка 40 м/с, а длительность спайка ПД— около 1,5 мс, то длина волны возбуждения будет 60 мм.
Любой периферический нерв содержит нервные волокна, отличающиеся друг от друга диаметром и толщиной миелиновой оболочки. Оба эти фактора существенно влияют на скорость проведения возбуждения. Скорость проведения возбуждения по миелинизированному волокну (волокно с толстой миелиновой оболочкой) определяется расстоянием между перехватами Ранвье: чем больше межперехватный участок, тем выше скорость проведения. При большем диаметре волокна затухание в этом участке уменьшается, поэтому длина межперехватного участка может быть больше. Кроме того, миелиновая оболочка у волокон с большим диаметром толще и утечка тока в связи с этим меньше. Благодаря этому у миелинизированных волокон большого диаметра длина межперехватного участка больше, надежность передачи импульса при этом сохраняется. Скорость проведения возбуждения пропорциональна диаметру волокна, а не корню из диаметра, как у безмякотных волокон.
Если регистрировать внеклеточно ПД седалищного нерва лягушки-быка и располагать отводящие электроды на расстоянии 150 мм от раздражающих, тогда получим ПД сложной формы: кроме высокоамплитудного спайка ПД имел ряд дополнительных колебаний (рис. 10). Этот факт обясняется наличием в нерве групп волокон разного диаметра, обладающих различной скоростью проведения, и разделили волокна седалищного нерва на три большие группы: А — миелинизированные волокна с диаметром 2,5-16 мкм и скоростью проведения 15-90 м/с; В — миелинизированные вегетативные волокна с диаметром около 3 мкм и скоростью проведения 10-15 м/с и С — безмякотные волокна с диаметром не более 2 мкм и скоростью 1-2 м/с. Группа А в свою очередь может быть разделена на три подгруппы: альфа, бета и гамма. Величины скорости проведения полученные на седалищном нерве травяной лягушки составляют: альфа -волокна - 42 м/с, бета - 25 м/с, гамма - 16 м/с.
Рис, 10. Г'рафик суммарного ПД нервного ствола (А) и его дисперсия (Б)
Работа 1. РЕГИСТРАЦИЯ ПОТЕНЦИАЛА ДЕЙСТВИЯ (ПД) СЕДАЛИЩНОГО НЕРВА ЛЯГУШКИ
Цель работы: Регистрация монофазного и двухфазного потенциала действия седалищного нерва лягушки. Измерение его амплитуды, длительности и времени деполяризации — реполяризации.
Потенциал действия (ПД) нервного ствола создается за счет алгебраической суммации ПД отдельных нервных волокон, образующих нерв. По своим свойствам ПД нерва во многом отличается от ПД отдельных волокон. Он не подчиняется закону "все или ничего", а, напротив, проявляет градуальную зависимость от силы стимула и т. д.
Возбужденный участок электроотрицателен по отношению к невозбужденному. Участок нерва, который оказывается в каждый данный момент электроотрицательным, имеет различную длину у разных нервов — от нескольких миллиметров до нескольких сантиметров — и соответствует длине волны. Отрицательный заряд распространяется вдоль всего нерва, причем длина волны остается постоянной. После прохождения нервного импульса электрический потенциал данного участка возвращается к исходной величине.

Рис. 11. Схема образования двухфазного ПД из двух монофазных
Когда нервный импульс находится вне участка, на котором расположены электроды, связанные с электронно-лучевым осциллографом (рис. 11), отклонение не регистрируется, так как между электродами О, и О2 нет разности потенциалов. Когда нервный импульс достигает электрода О] (рис. 11,2), расположенный под ним участок становится электроотрицательным. Между электродами О; и О2; возникает разность потенциалов, и осциллограф регистрирует отклонение (рис. 11,5) показывает момент нахождения импульса между электродами. При этом ткань, расположенная под обоими электродами, имеет одинаковый потенциал и, следовательно, отклонение не регистрируется. Когда волна отрицательного потенциала достигает электрода О2 (рис. II, 4\ возникают условия, сходные с теми, которые изображены на (рис 11, 2). Разница лишь в том, что теперь отклонение луча осциллографа имеет обратное направление, так как отрицательный потенциал расположен под вторым электродом.
Если представить потенциал как функцию времени, то получим кривую, изображенную на (рис. 11, 5) Так как отрицательный потенциал в определенной точке возникает и исчезает не мгновенно (для достижения максимума требуется небольшой интервал времени), то восходящий и нисходящий участки кривой имеют некоторый наклон. Для пояснения был приведен пример, который редко встречается в действительности. Обычно длина волны гораздо больше расстояния между электродами. Очевидно, что в таком случае между отклонениями, которые соответствуют появлению отрицательного потенциала под электродами О] и О3, не будет изо потенциальной линии. В тот момент, когда волна отрицательного заряда достигнет второго электрода, под первым электродом она еще не успеет исчезнуть полностью и получается кривая, показанная на (рис.11, б). Аналогичные рассуждения приводят к заключению, что амплитуда отклонения тем меньше, чем меньше расстояние между электродами.
Потенциал действия, зарегистрированный таким способом, называют двухфазным. Отрицательный потенциал под электродом, расположенным ближе к месту раздражения, регистрируется в виде отклонения луча осциллографа вверх, а под вторым электродом (более удаленным) — в виде отклонения луча вниз.
 
|
При изучении изменений потенциала удобнее регистрировать так называемый монофазный потенциал действия (рис. 12).
Рис. 12. Схема возникновения монофазного ПД
При такой регистрации предотвращается подход импульса ко второму электроду и записывается лишь прохождение импульса под первым электродом. Это можно осуществить различными способами. В простейшем случае нерв пережимают пинцетом между электродами или помещают электрод в поврежденный конец нерва. Наиболее удобным и t эффективным способом является деполяризация конечного участка нерва с помощью изотонического раствора КС1.
Для работы необходимы, установка для отведения потенциалов действия (усилитель биопотенциалов УБП, осциллограф С1-49, стимулятор, генератор парных импульсов Г5-26 с изолирующим трансформатором), стимулирующие и отводящие электроды, влажная камера, набор препаровальных инструментов, раствор Рингера, лягушка.
Проведение работы: Ком мутируют установку, как показано на рис. 13.
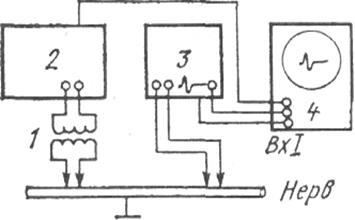
Рис.13. Блок-схема установки для регистрации ПД нервного ствола:
1 — изолирующая приставка. 2 — генератор парных импульсов Г5-26,
3 —УБП, 4 — осциллограф С1 -49
Готовят препарат: седалищный нерв — большеберцовый нерв (см. "Малый практикум по физиологии"). Помещают препарат во влажную камеру на стимулирующие и отводящие электроды так. чтобы расстояние между ними составляло 2-3 см. Раздражающие электроды подключают к выходу стимулятора, а отводящие соединяют с входом усилителя.
Ограничитель полосы частот усилителя сверху ставят на отметку "10 кГц", а ограничитель полосы снизу — на отметку "10 Гц". Переключатель хода работ усилителя устанавливают в положение "Работа". Ручки ступенчатой и плавной регулировки усиления ставят в среднее положение.
Устанавливают переключатель рода работ стимулятора в положение "Внутр. запуск", а регулятор частоты раздражения — в положение 5-20 имп/сек (Гц). Осциллограф переводят в ждущий режим работы с внешним запуском от синхроимпульса стимулятора. Время развертки подбирают в процессе работы.
Увеличивая постепенно от минимальной амплитуду прямоугольных импульсов стимулятора, наблюдают на экране осциллографа форму регистрируемой кривой. Сначала на экране возникает артефакт раздражения, то есть петля раздражающего тока, который достигает отводящих электродов за счет физического распространения тока. Артефакт отмечает момент раздражения нерва.
При дальнейшем повышения напряжения раздражающего тока артефакт увеличивается, и на изолинии появляется небольшая волна; волна возрастает до тех пор, пока не возникает выраженный потенциал действия (рис. 14).

Рис.14. Изменение ПД в ответ на увеличении амплитуды стимула
Регистрируют перечисленные этапы появления потенциала действия на кальке, накладывая ее на экран осциллографа и срисовывая кривые, или фотографируют их. Измеряют по шкале осциллографа время фазы деполяризации, реполяризации и длительность ПД.
Ставят переключатель рода работ усилителя в положение "калибровка" и выставляют калибровочный импульс (0,25-ь1 мВ), измеряя его на экране осциллографа. Сравнивают величину калибровочного сигнала с величиной потенциала действия и высчитывают его значение в милливольтах (определяют коэффициент усиления К = Uвых/Uвх) определяют амплитуду ПД
Инактивируют участок нерва под вторым отводящим электродом, положив на него ватный шарик, смоченный спиртом. Наблюдают постепенное исчезновение отрицательного колена потенциала действия, вплоть до появления монофазного ПД.
Для доказательства неподчинения потенциала действия нервного ствола, состоящего из большого числа нервных волокон различного диаметра, с различным порогом раздражения, закону "все или ничего", продолжают увеличивать амплитуду раздражения. Наблюдают, как по мере повышения силы раздражения происходит дальнейшее возрастание амплитуды потенциала действия. После того, как возбудятся все волокна, амплитуда ПД, несмотря на дальнейшее усиление раздражения (это можно видеть на экране по увеличению артефакта), больше не увеличивается (рис. 14).
Работа 2. ОПРЕДЕЛЕНИЕ ЗАВИСИМОСТИ СИЛА - ДЛИТЕЛЬНОСТЬ РАЗДРАЖЕНИЯ НЕРВА. ОПРЕДЕЛЕНИЕ ХРОНАКСИИ НЕРВА
Цель работы'. Определить зависимость между силой раздражающего тока и минимальной длительностью его действия, необходимой для возникновения возбуждения. Измерить хронаксию седалищного нерва лягушки.
Нервный импульс может возникать при действии различных химических, механических и электрических раздражений. В организме нервные импульсы возникают в результате раздражения различных рецепторов или возбуждающей активности нейронов.
Практически для искусственного раздражения наиболее удобны электрические импульсы, поскольку их легко охарактеризовать.
Естественные или искусственные импульсы вызывают изменение свойств мембраны нерва, изменяя главным образом ее проницаемость для различных ионов.
Пороговая сила раздражающего тока зависит от длительности его действия. Эта зависимость характерна для различных возбудимых тканей и описывается кривой, которая по существу представляет собой гиперболу. Сила тока и длительность его действия обратно пропорциональны. Чем короче раздражающий импульс, тем больше сила тока, необходимая для возникновения импульса. Асимптотическое приближение гиперболы к оси времени указывает на существование пороговой силы, ниже которой импульс не может быть вызван независимо от длительности раздражения. Эта величина получила название реобазы. Наиболее короткий интервал времени, в течение которого должен действовать ток, равный по величине реобазе, чтобы вызвать возбуждение, называют временем утилизации или полезным временем. Асимптотическое приближение гиперболы к оси ординат означает, что для возникновения импульса необходимо некоторое минимальное время действия раздражителя.
Кривую сила-длительность можно построить путем определения координат по оси времени для различных координат по оси тока или в обратном порядке. Поскольку измерения наиболее точны в средней части кривой, было предложено пользоваться хронаксией. Хронаксия — это время, в течение которого должен действовать ток, равный по силе удвоенной реобазе, чтобы вызвать возбуждение. Для измерения хронак-сии определяют минимальную длительность раздражения, равного по силе удвоенной реобазе. Реобаза — это пороговая сила продолжительного действия тока.
С помощью хронаксии можно сравнить возбудимость различных тканей.
Для работы необходимы: Аппаратура и вспомогательные материалы, как и для работы 1.
Проведение работы. Препарат седалищного нерва лягушки помещают в камеру и обычным способом подводят раздражающие и отводящие электроды. Определяют пороговую силу для раздражающего импульса большой длительности (60-И00 мс).
Появление небольшой активности на экране осциллографа характеризует пороговую величину раздражения (этого добиваются регулировкой амплитуды раздражения).
Постепенно укорачивают длительность импульса стимулятора и для каждого интервала времени определяют его пороговую силу. Пороговой называется такая сила раздражающего тока, которая как раз достаточна для возникновения активности. Силу (в вольтах, милливольтах) и длительность (в миллисекундах) импульса для каждой точки определяют по положению соответствующей шкалы регуляторов стимулятора.
Полученные парные величины наносят на миллиметровую бумагу (лучше полулогарифмическую) и строят кривую сила-длительность.
По этой кривой графически определяют хронаксию (X). Хронаксию можно определить и непосредственно в опыте. Для этого подают на нерв раздражающий импульс длительностью 60-=-100 мс и измеряют порог. Затем амплитуду импульса доводят до удвоенной величины порога (удвоенная реобаза). На экране осциллографа регистрируют при этом высокий потенциал действия. Укорачивают длительность раздражающего тока и определяют величину, при которой потенциал действия исчезает. Такая длительность импульса, характеризуемая определенным интервалом времени, и есть хронаксия. Записывают показания шкалы стимулятора в протокол.
Полученная таким образом величина хронаксии должна совпасть с величиной, полученной графически.
Работа 3. РЕФРАКТЕРНЫЙ ПЕРИОД НЕРВА
Цель работы: Определение абсолютного, относительного и супернормального рефрактерных периодов седалищного нерва лягушки.
Во время прохождения импульса по нервному волокну внутрь аксона входит некоторое количество натрия и выходит соответствующее количество калия. Эти количества малы и распределение ионов на обеих сторонах мембраны меняется весьма незначительно, и через аксон могут проходить следующие импульсы. Тем не менее, после продолжительной активности первоначальный ионный состав должен быть восстановлен. Это достигается в период покоя за счет активного переноса ионов против концентрационного градиента с помощью "натрий -калиевого насоса". Этот процесс и составляет, в основном, восстановительный цикл нервного волокна.
В ограниченном смысле под восстановительным циклом понимают изменения, которые происходят в мембране непосредственно после окончания ПД и продолжаются в течение десятых долей миллисекунды.
Сразу же после стимула, вызывающего нервный импульс, нервное волокно становиться рефрактерным (рис. 9) или возбудимым, по отношению к последующим стимулам, как бы велика не была их амплитуда. Этот промежуток времени называют абсолютным рефрактерным периодом. За ним следует фаза пониженной возбудимости, во время которой нервный импульс можно вызвать только при необычно большой силе раздражения, а амплитуда возникающего потенциала действия ниже, чем обычно. Это относительный рефрактерный период. Далее следует фаза, во время которой порог раздражения ниже, чем в норме, ее называют периодом супернормальной возбудимости.
По истечении названных периодов проницаемость мембраны для ионов нормализуется, и она вновь становиться способной генерировать импульс.
С физиологической точки зрения возбудимую ткань можно охарактеризовать по ее способности воспроизводить максимальное количество ответов в единицу времени. Это максимальная частота указывает на изменения возбудимости после нервного импульса.
Нерв раздражают импульсом, максимальным для наиболее лабильных волокон (группа А-волокон). Этот импульс называют кондиционирующим. Через различные интервалы времени нерв возбуждают повторным — тестирующим — импульсом различной силы.
Если интервал времени между двумя импульсами (кондиционирующим и тестирующим) достаточно велик, то при одной и той же силе импульсов регистрируются потенциалы действия с одинаковой амплитудой. Если интервал между двумя раздражающими импульсами короче относительного рефрактерного периода, то происходит уменьшение амплитуды ПД, возникающего в ответ на тестирующий стимул. Когда тестирующий импульс попадает в абсолютный рефрактерный период, то потенциал действия вообще не возникает.
Для работы необходимы'. Стимулятор парных импульсов, независимые друг от друга по амплитуде, и позволяющие регулировать интервал между двумя импульсами. Осциллограф С1-49, усилитель УБП, влажная камера, набор препаровальных инструментов, лягушка.
Проведение работы. Коммутируют установку как показано на рис. 13. Отпрепарированный седалищный нерв лягушки помещают во влажную камеру на раздражающие и отводящие электроды.
Устанавливают регуляторы регулировки длительности кондиционирующего и тестирующего импульсов стимулятора на равные значения (0,1-И мс) Частота импульсов подбирается экспериментально с целью фиксирования ПД на экране осциллографа (5-5-20 Гц)
Интервал времени между импульсами (задержка) делают порядка 7-^20 мсек. Регистрируют ПД.
Не меняя силы раздражения, постепенно уменьшают интервал между двумя импульсами, вращая регулятор задержки 2-го импульса стимулятора.
Начиная с определенного интервала времени между стимулами, амплитуда ПД, возникающая в ответ на тестирующий импульс, начинает уменьшаться. Это одна граница (конец) относительного рефрактерного периода. В конце концов (при дальнейшем уменьшении интервала между стимулами) второй ПД исчезает совсем. Этот момент ограничивает начало относительного рефрактерного периода.
Зарисовывают получаемые потенциалы действия (и первый, и второй), прикладывая кальку к экрану осциллографа (6-7 положений). Количественно периоды определяют, высчитывая интервалы времени между стимулами по шкале "задержка" стимулятора для каждого положения ПД на экране осциллографа.
Для доказательства существования относительного рефрактерного периода опыт повторяют, но на этот раз при одновременном уменьшении интервала между двумя стимулами, повышают силу тестирующего раздражения. Наблюдают возрастание амплитуды второго ПД до уровня первого.
Существование периода супернормальной возбудимости можно продемонстрировать следующим образом. Находят пороговую силу тестирующего раздражения при заведомо большом интервале времени между двумя импульсами. Затем уменьшают интервалы не меняя силы импульсов. При определенном интервале пороговое тестирующее раздражение становиться надпороговым.
Работа4. ОПРЕДЕЛЕНИЕ СКОРОСТИ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ ПО НЕРВУ
Цель работы: Определить в эксперименте скорость проведения возбуждения по седалищному нерву лягушки.
Общепринято, что распространение нервного импульса вдоль нервного волокна обеспечивается способностью возбужденной зоны стимулировать соседние области. Активная точка деполяризована и заряжена отрицательно по отношению к невозбужденным участкам. В результате возникает разность потенциалов между такой точкой и соседней областью. Разность потенциалов порождает местный ток, который течет от интактной поверхности по направлению к деполяризованному участку, и, таким образом, прогрессивно деполяризует соседнюю неактивную область.
Разность потенциалов, вызванная потенциалом действия, в несколько раз превышает величину, необходимую для возникновения нервного импульса в соседнем участке. Таким образом, существует большой "гарантийный" фактор, который обеспечивает проведение даже в неблагоприятных условиях, например, в период относительной рефрактерной фазы после развития нервного импульса.
Седалищный нерв лягушки — смешанный нерв. Он содержит волокна диаметром от 3 до 29 микрон. Поэтому при регистрации ПД получается плавная кривая, представляющая собой сумму сотен потенциалов действия от сотен волокон, проводящих импульсы с различной скоростью. В настоящей работе представляется возможным, т.о., определить суммарную скорость проведения по нерву (в основном, группы А-волокон).
Для работы необходимы: Установка для отведения ПД и вспомогательные инструменты, как и для предыдущей работы.
Проведение работы: Коммутируют установку, как показано на рис. 13. Готовят препарат седалищного нерва лягушки и помещают его на стимулирующие и отводящие электроды. Желательно, чтобы расстояние между ними (S) было не менее 30-^50 мм. Необходимыми условиями является фиксация стимулирующих и отводящих электродов на определенном расстоянии друг от друга. В процессе опыта это расстояние не должно меняться.
Переводят приборы в требуемый режим работы (см. предыдущую работу). Подбирают силу и скорость развертки раздражения так, чтобы на экране осциллографа наблюдалась картина, подобная той, которая приведена на рис. 14.
Для определения скорости проведения импульса необходимо знать путь, который проходит нервный импульс и время, затраченное на прохождение этого пути.
Путь прохождения нервного импульса "S" высчитывают, измеряя циркулем и линейкой расстояние от первого раздражающего до первого отводящего электродов, на которых лежит нерв.
Измеряют по шкале осциллографа время (Т) в мс, как отрезок линии от артефакта раздражения до восходящего колена ПД. Рассчитывают по формуле скорость проведения ПД
V - S/T (м/с).
РЕКОМЕНДУЕМАЯ ЛИТЕРАТУРА
1. Буреш Я., Петрань М., Захар И. Электрофизиологические методы исследвания. М: ИЛ. 1962.
2. Каталымов Л, Л., Сотников О.С. Физиология нейрона. Ульяновск. 1991.
3. Катц Б, Нерв, мышца и синапс. М.: Мир, 1968.
4. Шмидт Р., Тевс Г, Физиология человека. М.: Мир, 1996.
5. ЭккертР., РенделлД., ОгастинДж, Физиология животных. М.: Мир, 1991.
ОГЛАВЛЕНИЕ
Вводные замечания 3
1................................................................ Электронная аппаратура 3
1.1.Стимуляторы 3
Электроды 5
Усилители 10
Регистрирующая аппаратура 16
Электронные приборы специального назначения 20
2................................................................ Электрические свойства возбудимых тканей 21
Работа I. Регистрация потенциала (ПД) седалищного нерва лягушки 26
Работа 2. Определение зависимости сила-длительности раздра
жения нерва. Определение хронаксии нерва 3 1
Работа 3. Рефрактерный период нерва 33
Работа 4. Определение скорости проведения возбуждения по
нерву 35
Рекомендуемая литература 37