Другое возможное теперь направление исследования состоит в рассмотрении связи между исходным развитием нейронной цепи у эмбриона и ее последующей модификацией обучением. И при развитии, и при обучении в нервной системе происходят функциональные изменения — меняются эффективность синапсов и другие свойства нейронов. Как соотносятся такие изменения? Основаны ли механизмы обучения на механизмах пластичности, проявляющейся во время развития или же позднее формируются совершенно новые специализированные для обучения процессы?
Какими бы ни оказались ответы на эти интригующие вопросы, при исследовании беспозвоночных животных возникло поразительное и обнадеживающее обстоятельство: теперь стало возможным точно локализовать и наблюдать на клеточном - а в конце концов, возможно, и на молекулярном - уровне простые свойства памяти и обучения. Хотя для сложного головного мозга высших животных характерны некоторые высшие проявления умственной деятельности, теперь стало ясно, что элементарные свойства того, что считается мыслительными процессами, можно обнаружить в активности лишь очень малого числа нейронов. Поэтому и с философской, и с методической точек зрения интересно будет узнать, в какой степени высшие формы мышления можно объяснить действием более простых компонентов и механизмов. В той мере, в какой возможны такие редукционистские объяснения, важно также установить, как единицы этой элементарной азбуки комбинируются для создания языка гораздо более сложных мыслительных процессов.
У. НАУТА, М. ФЕЙРТАГ
Организация мозга
Головной и спинной мозг млекопитающих, включая человека, состоит из нескольких миллиардов нервных клеток, причем отдельные нейроны могут быть связаны с тысячами других. Как же организована эта огромная трехмерная сеть?
Мы видим два общих подхода к представлению о нейроанатомии. Первый — высокопарный: утверждается, что мозг — вместилище ума, чувств и желаний, памяти и способности учиться, и того любопытного ощущения, которое свойственно людям, — ощущения будущего. Затем мы начинаем внимательно рассматривать, как этот таинственный орган выглядит, так сказать, «во плоти». Определенные части мозга, в особенности кора больших полушарий, удивительным образом организованы; другие поражают своей кажущейся неупорядоченностью. Но даже и самые высокоупорядоченные структуры, в которых организация нейронов и разнообразных связей между ними напоминает электронные схемы, сопротивляются нашим сегодняшним попыткам понять их.

О порядке и беспорядке в клеточной организации мозговых структур можно судить по этим двум микрофотографиям. Обе они представляют тонкие срезы мозга кошки, которые подверглись двойному окрашиванию: по методу Гольджи, позволяющему выявить некоторые из нейронов со всеми их отростками в виде черных силуэтов, и по методу Ниссля, приводящему к окрашиванию всех клеточных тел в синий цвет. Метод Гольджи обеспечивает прокрашивание только 5 или менее процентов всех нейронов, по-видимому, по случайному выбору; если бы прокрашивались всенейроны, данная ткань выглядела бы однородно черной. А. Зубчатая извилина гиппокампа; тела нейронов в форме продолговатых пирамид располагаются в ряд, отсылая свои отростки почти параллельным пучком. Б. Микрофотография относится к крупноклеточной ретикулярной формации. Здесь нейроны образуют неупорядоченную сеть.
Второй подход более прозаический. Мозг просто подразделяется на части, поскольку подходящие методы окрашивания выявляют в одних местах скопления нейронов, опутанных густой сетью их же собственных нитевидных отростков, в других же ткань состоит в основном из длинных Волокон нервных клеток — аксонов, которые обслуживают связи на далекие расстояния в нервной системе. Ткань первого рода — это серое вещество, второго — белое вещество.
Конечно, есть искушение приписать каждой области какую-нибудь функцию, как если бы мозг в целом был чем-то вроде радиоприемника. Но существо работы центральной нервной системы - головного и спинного мозга — это проведение входящей сенсорной информации к множеству структур и конвергенция на нейронах, которые возбуждают эффекторные органы: мышцы и железы. Следовательно, система в целом обладает свойствами, выходящими за пределы тех, которые можно обнаружить у простого набора модулей.
Возьмем, например, такую структуру мозга как субталамическое ядро. Его разрушение у человека ведет к моторной дисфункции, известной под названием гемибаллизма, при которой больной делает непроизвольные движения, как бы бросая мяч. Следует ли отсюда, что нормальной функцией субталамического ядра должно быть подавление движений, напоминающих бросание мяча? Конечно, нет; данное состояние больного только дает представление о работе центральной нервной системы, выведенной из равновесия отсутствием субталамического ядра.
Мы упоминаем эти вещи, чтобы установить ограничения для любых объяснений анатомии мозга. Мы дадим здесь какое-то объяснение, но оно по необходимости будет несколько расплывчатым. Делать иного рода предположения было бы просто неправомерно.
Полезно сделать некоторые предварительные замечания. В первые десятилетия этого века Дж. Паркер (G. Parker) из Йельского университета искал самую примитивную рефлекторную дугу. Такие дуги были идентифицированы у позвоночных животных; это состоящие из одного или нескольких нейронов пути, с помощью которых возбуждение, порождаемое сенсорным стимулом, действующим на некоторую часть тела, может быть проведено к эффекторной ткани и, соответственно, вызвать движение. Во времена Паркера рефлекторные дуги рассматривались как простейшие схемы, посредством которых природа объединяет клетки в нервную систему; в соответствии с этим было широко распространено мнение, что нервная система возникает тогда, когда некоторый организм приходит к необходимости иметь какую-нибудь клетку или цепочку клеток для посредничества между внешним стимулом и ответным движением животного. Со временем предполагалось установить, что у более развитых организмов эволюция нервной системы продвигалась вперед путем увеличения числа и сложности таких цепочек.
Сначала внимание Паркера привлекли эпителиальные слои некоторых морских гидроидных полипов и актиний, потому что иногда они содержали клетки, которые выглядели (если были подходящим образом окрашены) как нейроны. У основания такой клетки Паркеру удалось разглядеть начало нити, весьма похожей на аксон, которая, подходя к мышечному волокну, разветвлялась. Паркер не был уверен в том, что тут образовывался контакт, но предположил, что между аксоном и мышечным волокном устанавливается какая-то связь. Конечно, он был прав, но это устройство весьма примитивно; его схему можно было бы назвать однонейронной нервной системой, поскольку вся линия проведения состоит всего лишь из одной клетки. Что такая нервная система будет делать в ответ на стимул, столь же предсказуемо, как и то, что будет делать дверной звонок, если нажать на кнопку. В то же время относительно нервной системы человека ясно, что поведение, на которое она делает способными человеческие существа, самое что ни на есть непредсказуемое.
Очевидно, что-то должно вмешиваться в работу механизма дверного звонка, и потому Паркер исследовал ситуацию у несколько более сложных организмов. В эпителиальном слое у некоторых полипов и медуз он обнаружил нейронные структуры, сходные с найденными им ранее. Однако под эпителием он теперь нашел дополнительные нейроны, которые вместе образовали широко разветвляющуюся сеть. Таким образом, нервная система этой второй группы организмов оказалась усложненной: нейроны эпителиального слоя образовывали контакты с субэпителиальной сетью, а клетки этой сети в свою очередь образовывали контакты с сократимыми тканями в глубине организма. Следовательно, уже можно было говорить о двухнейронной нервной системе, в которой сенсорные нейроны (у этих простых созданий нейроны лежат вблизи поверхности и находятся в непосредственном контакте с окружающей средой) связаны с мотонейронами (нейронами, которые образуют контакты с эффектерными клетками, в данном случае — сократимыми клетками, и, значит, по существу, с мышечными волокнами).
Остается ли эта сеть в высшей степени предсказуемой? По-видимому, нет. Предположим, что мотонейроны связаны друг с другом и на вход каждого из них поступают не только сообщения, идущие из окружающей среды через посредство сенсорных нейронов, но и сообщения от соседних мотонейронов. Вообразим далее, что некоторые из этих сообщений могут быть возбуждающими, т.е. заставляют данный мотонейрон с большей готовностью генерировать и передавать свои собственные сигналы в ответ на поступление других сигналов, и что другие сообщения могут быть тормозными. В таких условиях приходится решать вот какую задачу: чтобы предсказать, что будет делать некоторый нейрон в ответ на различные входные сигналы, нужно алгебраически просуммировать возбуждающие и тормозные сообщения, которые на нем конвергируют.
Возможно, что такое двухнейронное устройство позволяет облагодетельствованной природой медузе быть более непредсказуемой в своем поведении, чем актинии и другие организмы с однонейронной нервной системой. Однако затем следует дальнейшее достижение, и оно тоже обнаружено у очень примитивных организмов — некоторых других меду:». В каком-то смысле это высшее достижение, так как и нервная система таких медуз, и нервная система чело века — обе состоят по существу только из трех классов нейронов. У этих медуз, как и у человека, сенсорныенейроны, как правило, больше не связаны непосредственно с мотонейронами. Между этими двумя элементами развивается барьер из нейронов, которые имеют взаимосвязи не только с мотонейронами, но и друг с другом.
Правда, этот третий и последний шаг, возможно, уже был сделан всеми организмами, которые имеют субэпителиальную сеть нервных клеток. Выше, при рассмотрении двухнейронной нервной системы, предполагалось, что все клетки, составляющие этот слой, — мотонейроны, т.е. клетки, иннервирующие эффекторные ткани. Однако в действительности такие связи могут устанавливать только некоторые из многих субэпителиальных клеток. Остальные могут располагаться в сплетении таким образом, что получают на вход сингалы от сенсорных нейронов эпителия, а сообщаются лишь с другими нейронами того же типа или с мотонейронами, но не с эффекторными структурами. Не являясь ни сенсорными, ни моторными, они служат посредниками при проведении сигнала по сенсомоторному пути.
Короче, здесь тоже имеются промежуточные (вставочные) нейроны. Хотя трехнейронную организацию трудно вычленить в диффузной нейронной сети, она широко представлена на более поздних стадиях эволюции; у животных, более высоко развитых, чем медузы, диффузная субэпителиальная нервная сеть концентрируется либо в последовательность сегментарных ганглиев (скоплений нейронов), либо в единую несегментированную центральную нервную систему. Таким образом, критическим моментом явилось изобретение «великой промежуточной сети» — барьера из промежуточных нейронов, которые вклинились между сенсорными нейронами и мотонейронами, на раннем этапе эволюции животных.
Насколько далеко зашел прогресс в развитии этой промежуточной сети на сегодняшний день, легче всего показать с помощью некоторых цифр. Начнем с вопроса о том, сколько нейронов в центральной нервной системе человека. В качестве ответа часто можно услышать: порядка 1010. Это результат подсчета вставочных нейронов и мотонейронов, ибо случилось так, что настоящие сенсорные нейроны лежат не в центральной нервной системе, а в ганглиях, которые расположены вне головного и спинного мозга. Это весьма привлекательная цифра, которую легко запоминать и использовать. И все же имеются еще классы нейронов, которые столь малы и столь плотно упакованы, что оценить их число трудно или невозможно. Один из таких классов — это клетки-зерна. Как раз в одной из частей головного мозга — мозжечке — находится так много клеток-зерен, что величина 1010нейронов для всей центральной нервной системы становится сомнительной. Общее число вполне может быть на порядок, а возможно, и на два порядка больше.
Все же примем на минуту, что общее число действительно 1010. Сколько же из этих клеток мотонейроны? Ответ таков, что их, оказывается, не может быть больше двух или трех миллионов. Эта цифра представляется смущающе малой в свете того факта, что только через мотонейроны работа нервной системы может выразиться в движении. Более того, такой ответ заставляет предполагать, что на мотонейроны должно конвергировать неправдоподобно большое число влияний; другими словами, это заставляет предполагать, что типичный мотонейрон должен образовывать синаптические связи с огромным числом аксонов, исходящих от равно огромного числа нейронов великой промежуточной сети.
Полагают, что на поверхности типичного мотонейрона в спинном мозгу человека имеется что-то около 10000 синаптических контактов, из которых около 2000 - на клеточном теле и 8000 - надендритах - отростках, ветвящихся локально, в отличие от единственного аксона. Это не означает, что на данный нейрон посягают 10000 вставочных нейронов: когда такие нейроны устанавливают связи с какой-то клеткой, они имеют тенденцию образовывать множественные синаптические контакты. Однако все равно получается, что средний мотонейрон должен подвергаться массированному воздействию: число 1010 для нейронов центральной нервной системы подразумевает, что на каждый мотонейрон приходится от 3000 до 5000 нейронов великой промежуточной сети.
Вот какое последнее заключение должно быть выведено из цифр, которые мы процитировали: весь головной и спинной мозг человека - это великая промежуточная сеть, за исключением явно немногих миллионов мотонейронов. А когда великая промежуточная сеть начинает включать 99,98 процента всех нейронов, составляющих центральную нервную систему, этот термин теряет большую часть своего смысла: он начинает отражать саму ту сложность, с которой каждый должен столкнуться при попытках постигнуть нервную систему. Этот термин остается полезным только как напоминание о том, что большая часть нейронов мозга не принадлежит ни к сенсорным, ни к моторным. Строго говоря, они вставлены между чисто сенсорной частью организма и чисто моторной его частью. Они являются компонентами вычислительной сети.
Вторая группа предварительных замечаний касается общей анатомии центральной нервной системы. Отметим, в частности, что у всех видов позвоночных головной и спинной мозг сначала появляются у зародыша всего лишь в виде трубки толщиной в один клеточный слой. В передней части этой нервной трубки, которая в конечном итоге будет заключена в череп, вскоре появляются три утолщения — первичные мозговые пузыри. Это ромбэнцефалон, или задний мозг; мезэнцефалон, или средний мозг; и прозэнцефалон, или передний мозг. («Энцефалон» происходит от греческого «внутри головы».)
Из этих трех первичных пузырей передний пузырь является самым продуктивным по числу образующихся отделов и дальнейшей дифференцировке. Основным событием его эмбрионального развития является образование камер с левой и правой стороны. Они превращаются в полушария большого мозга; эту часть мозга часто называют также конечным мозгом (телэнцефалон); у некоторых видов он имеет скромные размеры, у других - чрезвычайно большие. Между полушариями лежит непарный центральный отдел переднего мозга, от которого отпочковались полушария. Он имеет название промежуточный мозг, или диэнцефалон.

Эти два среза фиксированного препарата мозга человека иллюстрируют сложность внутреннего строения мозга. Срезы были обработаны краской, которая избирательно окрашивает в черный цвет жировую миелиновую оболочку нервных волокон; в результате белое вещество предстает как черное, а серое (состоящее преимущественно из клеточных тел) остается неокрашенным. Пустые пространства на этих срезах указывают положение желудочков—заполненных жидкостью полостей внутри мозга. Верхний рисунок. Срез проходит примерно посередине мозга и захватывает кору больших полушарий, гиппокамп и таламус. Нижний рисунок. Срез сдвинут кзади и проходит через ствол мозга. Как сделаны срезы и где располагаются упомянутые структуры, поясняет следующий рисунок. (Препарат из коллекции профессора П. Яковлева из Гарвардской медицинской школы.)

Мозг человека разрезан на пять частей и раскрыт как книжка, что позволяет проследить связи между наружным и внутренним строением. Два сечения, представленные на предыдущем рисунке, обозначены буквами А и Б.
В процессе развития из переднего мозга выделяется еще одна пара камер-глазные пузыри. Их имеют даже незрячие животные, а у животных, способных видеть, они удлиняются в направлении к поверхности головы и в конечном счете превращаются в две сетчатки, соединенные с основанием переднего мозга своими стеблями-зрительными нервами. Наконец, из внутренней поверхности первичного переднего мозга развивается непарная срединная камера, которая дифференцируется и образует заднюю долю гипофизарного комплекса.



Два метода окрашивания дают возможность получить взаимно дополняющие друг друга изображения внутреннего строения мозга крысы. А. Показано сечение, проходящее чуть в стороне от срединной плоскости параллельно ей; срез был обработан по методу Ниссля, который позволяет избирательно окрашивать клеточные тела. Следовательно, каждая точка на микрофотографии соответствует какой-то отдельной клетке. Б. Срез обработан по методу Лойе (Loyez), который позволяет избирательно окрашивать миелинизированные волокна, оставляя тела клеток непрокрашенными, благодаря чему выделяются приводящие пути. В. Схема, на которой показаны различные структуры мозга.
На нижнем рисунке на стр. 92 показан результат этого эмбрионального роста. Такая схема в общем выдерживается для всех млекопитающих; она изображает полностью сформированную центральную нервную систему млекопитающего, разбитую на несколько отделов. Слева - спинной мозг (на рисунке — сильно укороченный). Вправо от него, без какой-либо резкой границы, отходит задний мозг — самый нижний отдел головного мозга. Его дорсальная часть (часть, наиболее близкая к спине животного) - придаток, называемый мозжечком.
За задним мозгом идет средний мозг, который у млекопитающих включает две пары структур, область из четырех бугров, известную под названием четверохолмия, крыши среднего мозга, или тектума («тектум» в переводе — «крыша»). Нижняя пара структур называется нижним двухолмием, верхняя пара — верхним двухолмием. Кроме этого, средний мозг дает мало оснований для подразделения, по крайней мере в продольном направлении. В сущности это довольно короткий участок человеческого мозга.
Далее идет центральный непарный отдел переднего мозга — промежуточный мозг. Его дорсальные две трети — это таламус. Остальная часть — гипоталамус. (Несколько в стороне от гипоталамуса есть третий отдел промежуточного мозга — субталамус, самую удивительную группу клеток которого — субталамическое ядро — мы упоминали вначале. Включение его усложнило бы рисунок.) Гипоталамус характеризуется наличием железистого придатка, называемого гипофизарным комплексом. Он также продолжается в переднем направлении, переходя в перегородку — структуру, которую, несмотря на ее расположение, лучше всего классифицировать как принадлежащую к промежуточному мозгу. Остальная часть переднего мозга — конечный мозг, мозговые полушария. В мозгу млекопитающих это наибольшая часть, намного превосходящая другие, и у многих видов млекопитающих оболочка полушарий — мозговой плащ, или кора большого мозга, — испещрена извилинами (gyri) и бороздами (sulci). У основания каждого полушария имеется простирающаяся вперед структура, которая полностью состоит из серого вещества (коры), хотя это и кора с очень примитивной клеточной структурой. Ее вздутый передний конец — обонятельная луковица, а ее ножка — обонятельный стебель; только та часть коры, которая лежит непосредственно под остальной частью полушария, является настоящей обонятельной корой. Вторая большая часть коры мозга у млекопитающих находится там, где мозговой плащ подворачивается и образует сложные складки, которые в поперечном сечении напоминают орнамент в стиле рококо. Эта замечательная структура известна под названием гиппокампа.
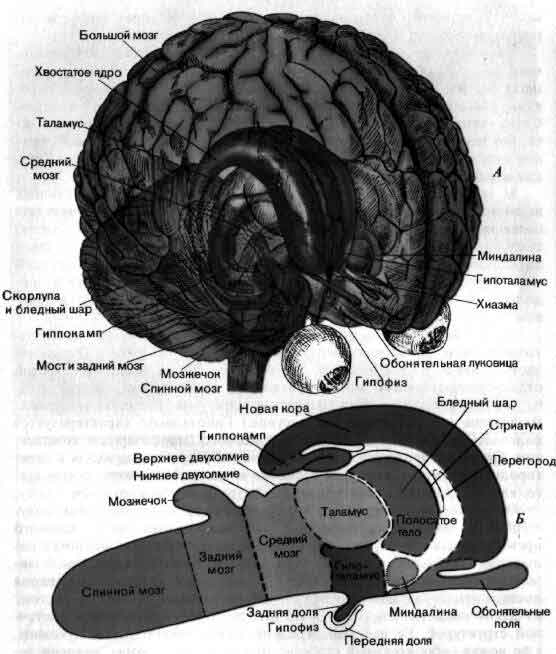
Головной и спинной мозг человека и других млекопитающих можно подразделить на отделы, исходя из данных анатомии, эмбриологии и клеточной организации. А. Мозг человека изображен таким образом, что его внутренние структуры просматриваются сквозь «прозрачные» наружные слои мозжечка. Б. Обобщенная схема мозга млекопитающего; эта схема условно принята для всех остальных рисунков данной статьи. Соответствующие структуры на обоих рисунках окрашены одинаково. Обычно принято делить мозг на передний, средний и задний. Задний мозг включает мозжечок. К среднему мозгу относятся бугорки четверохолмия - нижнее и верхнее двухолмия. Строение переднего мозга более сложно. Его наружная часть - полушарие, поверхность которого - складчатый пласт мозговой коры, включающей гиппокамп, новую кору и обонятельные поля. Внутри полушария располагаются миндалина и полосатое тело; последнее подразделяется на бледный шар и стриатум, включающий хвостатое ядро и чечевицеобразное ядро. Остальная часть переднего мозга — промежуточный мозг: верхние его две трети составляет таламус (который имеет множество подразделении), а нижнюю треть - гипоталамус (который соединяется с гипофизарным комплексом).
После выделения перечисленных частей остается еще одна часть коры большого мозга млекопитающих, которая имеет большую протяженность и чрезвычайно сложное строение; у человека и других приматов она, согласно оценкам, содержит не менее 70 процентов всех нейронов центральной нервной системы. Это новая кора — самая поздняя кора в процессе эволюции. Мы приобрели ее благодаря расхождению путей эволюции: после рептилий одна линия животных развивалась, усовершенствуя прежний тип, и дошла до птиц, а другая — более «авантюрная» - приобрела новую кору и дошла до млекопитающих. Таким образом, с чисто филогенетической точки зрения птицы воплощают в себе логическое завершение традиционного развития мозга, а млекопитающие представляют собой отклонение, поскольку они не имеют в своей родословной птиц. В результате одного из многочисленных разветвлений эволюции млекопитающих появились приматы - отряд, в котором новая кора достигает своего максимального развития. Мы, человеческие существа, пожинаем все плоды этого развития, возможно, включая и психопатологию.
В глубине каждого полушария большого мозга млекопитающих имеется несколько скоплений серого вещества. Одно из них — это миндалина, которая лежит под обонятельной корой. Другое - полосатое тело, находящееся в самой середине полушария. Это тело в свою очередь подразделяется на две части, которые различаются по клеточному составу. Первая из них — это внутренняя зона, называемая палеостриатумом, или бледным шаром. Вторая зона - наружная; она известна под названием стриатум.
Вернемся теперь к схеме связей в центральной нервной системе млекопитающих. Начнем с идентификации сенсорных нейронов, типа тех, которые Паркер нашел в эпителиальном слое медуз. Однако у позвоночных расположение сенсорных нейронов совсем иное. Известен всего один пример, где сенсорный нейрон является также рецептором на поверхности тела: только обонятельные эпителиальные клетки в слизистой крыши носа контактируют с внешней средой. Все другие сенсорные нейроны в теле позвоночных надежно упрятаны под поверхность и находятся в ганглиях, располагающихся вдоль спинного мозга по всей его длине, или в аналогичных ганглиях, лежащих в стороне от мозга. (У позвоночных термин «ганглий» сохранен для скоплений нейронов вне центральной нервной системы.) У каждого сенсорного нейрона есть аксон, который делится на две части: одна часть идет в центральную нервную систему, а другая иннервирует периферические структуры.

Соматосенсорная информация, например сообщения о кожных ощущениях, передается по нескольким путям спинного мозга. В левой части схемы показана пара репрезентативных сенсорныхнейронов, доставляющих сигналы от периферических сенсорных рецепторов в спинной мозг. Отсюда один путь сразу же ответвляется к мотонейронам (сплошные треугольники), волокна которых идут за пределы мозга к скелетным мышцам. Все другие пути сначала идут в скопления вторичных сенсорных нейронов, располагающиеся либо в той же самой области спинного мозга, либо в его верхней части, в ядрах дорсального столба. Путь, называемый медиальным лемниском, поднимается вверх от ядер дорсального столба к вентральному ядру таламуса, которое в свою очередь посылает волокна в соматосенсорную зону коры. Второй путь, называемый спино-таламическим трактом, поднимается к переднему мозгу из скоплений вторичных сенсорных клеток по всей длине спинного мозга, по ходу раздавая в стороны часть своих волокон. Малая доля волокон спино-таламического тракта в конечном счете достигает вентрального ядра. Скопления вторичных сенсорных клеток также посылают свои волокна в мозжечок.
На приведенном здесь рисунке, одна из таких клеток — назовем ее первичным сенсорным нейроном — посылает свой аксон в спинной мозг, доставляя туда сенсорные сообщения о таких событиях, как прикосновение к коже, движение сустава или сокращение мышцы. Эти сообщения не сразу поступают на мотонейроны; свои первые синаптические контакты сенсорный нейронустанавливает с так называемыми вставочными нейронами.
Однако имеется одно исключение. Это моносинаптическая рефлекторная дуга, образуемая боковой ветвью первичного сенсорного волокна, идущей «напролом» и устанавливающей прямой синаптический контакт с мотонейроном. Сначала это кажется обескураживающим: всего лишь несколькими абзацами выше мы отмечали, что после самых ранних ступеней эволюции нервной системы мотонейроны больше не утруждают себя обработкой сырых данных. Мы предположили, что вместо этого они теперь получают краткий итог обработки информации («дайджест»)нейронами великой промежуточной сети. Следовательно, моносинаптическая дуга может рассматриваться как очень примитивный тип нервной сети; с другой стороны, ее можно считать относительно новой: по-видимому, ее вновь открыли только наземные животные. В конце концов, воздух и земля — самые суровые среды; для горного козла один неверный шаг может оказаться роковым. В противоположность этому рыба может без всякого риска для себя сделать любое количество аналогичных ошибочных движений. Рыбы прекрасно уравновешены в воде, и сила тяжести для них далеко не так обременительна и враждебна. Таким образом, именно наземное, а не водное существование, по-видимому, требует наличия высоконадежных рефлекторных систем для удержания равновесия и, в частности, наличия пути, по которому мышца может сигнализировать соответствующим мотонейронам (и только им), что она чрезмерно растянута силой тяжести.
Моносинаптические рефлекторные дуги никогда не обнаруживаются за пределами таксах корректирующих механизмов. Так что короткие цепи между сенсорным входом и моторным выходом оказываются представленными в ничтожно малом числе. Подавляющее большинство первичных сенсорных волокон млекопитающих входит в великую промежуточную сеть и синаптически контактирует с клетками, составляющими группу, которую мы будем называть группой вторичных сенсорных клеток: это нейроны, стоящие первыми в цепи приема первичных сенсорных сигналов. Оттуда многие пути более или менее прямо направляются к мотонейронам. Все они в совокупности могут быть названы локальным рефлекторным путем, если не упускать из виду, что слово «локальный» может ввести в заблуждение, так как есть несколько рефлексов, которые вовлекают в работу весь спинной мозг по всей его длине, но тем не менее относятся к локальным, поскольку не выходят за его пределы. Первое звено в локальном рефлекторном пути — это некоторый нейрон из группы вторичных сенсорных клеток. Многие из таких клеток сами не образуют контактов с мотонейронами; вместо этого они синаптически переключаются на некоторые другие нейроны великой промежуточной сети, и только уже эти последние нейроны, наконец, замыкают дугу.
Другие пути состоят из аксонов, не идущих к мотонейронам. Возьмем мозжечковые каналы: из скоплений вторичных сенсорных нейронов заднего и спинного мозга многие аксоны восходят прямо к мозжечку. Показанный на рисунке на стр. 94 аксон, который идет таким образом, берет свое начало в группе вторичных сенсорных клеток спинного мозга и потому называется спино-мозжечковым волокном. (Слова «аксон» и «волокно» в нейроанатомии используются как синонимы.)
Третий путь — это лемнисковый путь. Слово «лемниск» в переводе с латинского — лента (петля); здесь им обозначаются пучки волокон, которые берут начало в группах вторичных сенсорных клеток и поднимаются к переднему мозгу, в частности к таламусу. На рисунке (стр. 94) показано, как один такой пучок идет по центру спинного мозга. В действительности он проходитлатеральнее: упрощенную иллюстративную схему трудно сделать топографически точной. Этот пучок назван спино-таламическим трактом, хотя только одно из трех его репрезентативных волокон изображено как оканчивающееся в таламусе. Два других волокна сопровождают его на некотором расстоянии, а затем, так сказать, «совершают непредвиденную посадку»: на рисунке показано, что оба оканчиваются на нейронах заднего мозга, хотя с таким же успехом они могли бы окончиться несколько далее по ходу — в среднем мозгу. Важно то, что из волокон спино-таламического тракта лишь небольшая доля действительно достигает таламуса. Тем не менее тракт назван в честь этого преуспевшего меньшинства, которое оканчивается в специфической части таламуса — вентральном ядре. Здесь волокна образуют синапсы с таламическими нейронами, аксоны которых идут, не прерываясь, в специализированное поле новой коры, называемое соматосенсорной корой.