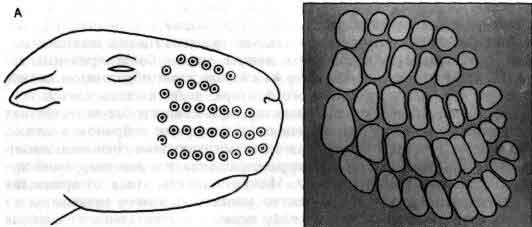

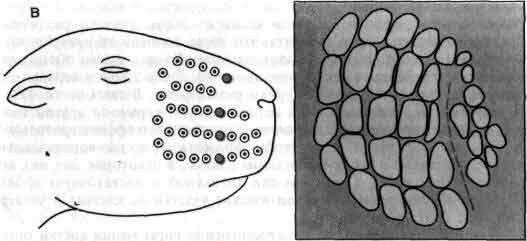
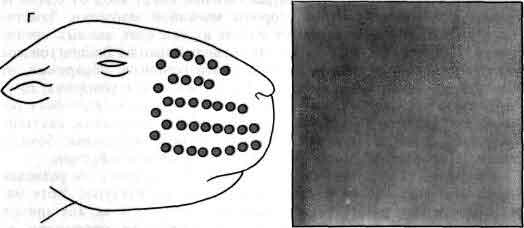
Вибриссы и бочонки соматосенсорной коры молодых мышей служат примером одной из многих систем, в которых обнаружена определяющая их зависимость топографии развивающейся нервной системы от ее входов. Вибриссы - это чувствительные волоски на мордочке мыши, а бочонки - это специализированные скопления нейронов в слое IV коры головного мозга. Входом для каждого бочонка служит одиночная вибрисса с контралатеральной стороны мордочки (А). Если вскоре после рождения животного разрушить один из рядов вибрисс, то позднее будет обнаружено, что соответствующий ряд бочонков в коре большого мозга отсутствует, а соседние бочонки увеличены (Б, В). Если разрушить все вибриссы, то бочонки вовсе исчезнут (Г). Должна иметь место значительная пластичность в развивающейся коре, поскольку волокна, иннервирующие вибриссы, не прямо контактируют с корой, а связаны с ней посредством по крайней мере двух синаптических переключений. Рисунок основан на работе Т. Вулси из Медицинской школы Вашингтонского университета.
У макака информация из сетчатки достигает четвертого слоя зрительной коры через посредство структуры, названной латеральным коленчатым телом. На этом уровне входы в кору от каждого глаза совершенно раздельны, что было прямыми методами показано на подопытных животных с помощью инъекций большого количества меченых аминокислот в одно глазное яблоко. Ганглиозные клетки сетчатки поглощают аминокислоту, включают ее в белок и транспортируют в латеральное коленчатое тело. Здесь некоторая часть метки освобождается и становится доступной для включения в клетки коленчатого тела, которые транспортируют ее по своим аксонам далее в зрительную кору. На соответствующим образом приготовленных радиоавтографах (которые позволяют выявить распределение меченых волокон, достигающих коры) видно, что первичная зрительная область организована в виде перемежающихся так называемых глазодоминантных полосок, причем каждая полоса имеет ширину около 400 мкм и получает входные сигналы либо от правого, либо от левого глаза. Д. Хьюбел, Т. Визель и С. Ле Вэй (D. Hubel, Т. Wiesel, S. Le Vay) из Гарвардской медицинской школы показали, что если сшить веки одного глаза у подопытного новорожденного животного (так чтобы сетчатка глаза не подвергалась воздействию структурированных зрительных стимулов), глазодоминантные полоски, связанные с закрытым глазом, будут заметно уже, чем в норме. В то же время полоски проекции открытого глаза соответственно расширятся (общая ширина двух соседних полосок, различающихся стороной стимуляции, при этом постоянна).
Результат этот частично вызван сжатием глазо-доминантных полосок «выключенного» глаза, сопровождающимся вторичным расширением полосок нормального глаза, и отчасти - сохранением неплотной группировки волокон из нормального глаза, имевшей место на более ранней стадии. Если исследовать входы от обоих глаз на различных стадиях развития, можно показать, что когда волокна из латерального коленчатого тела только достигают зрительной коры, входы обоих глаз существенно перекрываются в зоне проекции. Лишь к концу первого месяца жизни животного полоски четко разделяются. В свете этого факта (и результатов экспериментов, в которых затем закрывали другой глаз, и открывали первый) представляется вероятным, что эффект зрительнойдепривации состоит в том, чтобы привести геникуло-кортикальные клетки, связанные с нестимулируемым глазом, в некоторое для них невыгодное положение, в котором они проявляют недостаточную эффективность в конкуренции за синаптические участки на клетках в четвертом слое зрительной коры.
В соответствующем слое соматосенсорной коры мыши клетки организованы в четкие группы, названные бочонками. Физиологические исследования показывают, что каждый бочонок имеет вход от одной вибриссы с контралатеральной стороны мышиной мордочки. Заметим, что у мыши вибриссы являются одним из наиболее важных органов чувств. Т. Вулси (Th. Woolsey) из Медицинской школы Вашингтонского университета, первым открывший значение бочонков, обнаружил, что если удалить небольшую группу вибрисс сразу после рождения, то соответствующая группа бочонков в коре не разовьется. Этот факт особенно интересен потому, что между сенсорными нейронами, связанными с вибриссами, и нейронами, образующими кортикальные бочонки, лежат по крайней мере еще две группы вставочных нейронов.
Эти и многие другие наблюдения приводят к выводу, что развивающийся мозг является исключительно пластичной структурой. Хотя многие области могут быть «жестко запаяны», другие (такие, как зрительная кора) открыты для различных воздействий - как внутренних, так и внешних. Способность мозга к реорганизации в ответ на внешние воздействия или на локальное повреждение в настоящее время является наиболее активно изучаемым явлением нейробиологии не только потому, что очевидна его связь с такими феноменами, как обучение и память, и его отношение к способности мозга восстанавливаться после повреждения, но и потому, что они открывают тайны нормального развития мозга.
В заключение хочется подчеркнуть, что развитие мозга, как и развитие большинства других биологических структур, не обходится без ошибок. Ранее уже было упомянуто, что ошибки могут происходить во время миграции нейронов. Известны такие примеры возникновения ошибок во время формирования связей. В зрительной системе, как было замечено рядом исследователей, некоторые волокна зрительного нерва, которые должны были бы пересечь среднюю линию в составе перекреста, начинают в результате ошибки расти к ипсилатеральной стороне мозга. В некоторых таких ситуациях, если у животного на ранней стадии развития удалить один глаз, то число волокон, отклонившихся от правильного направления, может значительно возрасти. Поскольку аномально ориентированные волокна часто не обнаруживаются в зрелом мозгу, создается впечатление, что такие неправильно адресованные нейроны (и любые ими сформированные ошибочные связи) ликвидируются на более поздних стадиях развития. Остается загадкой, как выясняется, что они ошибочные, и каким образом они устраняются. Учитывая сложность механизмов развития, не приходится удивляться тому, что ошибки появляются. Удивительнее то, что они появляются редко и что чаще всего они эффективно исправляются.
Л. ИВЕРСЕН
Химия мозга
Сигналы передаются от нейрона к нейрону разными химическими медиаторами. Эти химические системы, наложенные на нейронные цепи головного мозга, добавляют к его функции еще одно измерение
Нейроны имеют биохимический аппарат, общий со всеми остальными живыми клетками, в том числе способность генерировать химическую энергию путем окисления пищевых веществ, а также восстанавливать и сохранять свою целостность. Нейроны обладают, кроме того, специфическими свойствами, которых лишены другие клетки и которые связаны с особой функциейнейронов как передатчиков нервных импульсов; сюда относятся необходимость в поддержании ионных градиентов, что требует большой затраты энергии, и свойства, связанные со способностью нейронов производить и выделять набор химических передатчиков, называемых нейромедиаторами. В синапсах - микроскопических участках, где тесно соприкасаются окончание одного нейрона и воспринимающая поверхность другого, приход импульса вызывает внезапное выделение молекул медиатора из, окончания. Затем эти молекулы диффундируют через заполненную жидкостью щель между двумя клетками и воздействуют на специфические рецепторы постсинаптической мембраны, изменяя при этом электрическую активность воспринимающего нейрона.
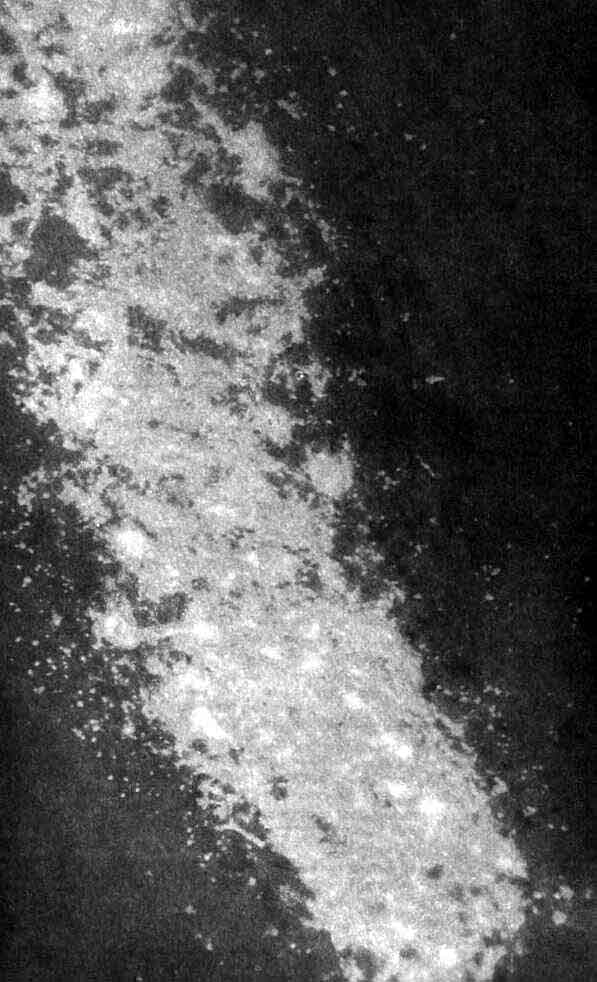
Нейроны, содержащие норадреналин - химический медиатор в головном мозгу, - ярко светятся на этом срезе мозга крысы под флуоресцентным микроскопом. Такие клетки, расположенные в участке мозга, именуемом locus coeruleus, стали видны под воздействием глиоксиловой кислоты, которая превращает норадреналин в его флуоресцирующее производное. В этом поле находятся еще тысячи других нейронов, но в них содержатся другие медиаторы и поэтому они не видны. Норадреналиновые нейроны в locus coeruleus посылают свои аксоны во многие отделы мозга, в том числе в мозжечок и передний мозг. Как полагают, они имеют отношение к регуляции сна, настроения, а также к системе поощрения. Микрофотография получена Ф. Блумом (F. Bloom), Г. Джонсом (G. Jones) и Ж. Мак-Джинти (J. McGinty) из Института Солка.

Химическая передача через синапс - узкую щель между двумя нейронами - в головном мозгу состоит из сложной последовательности молекулярных процессов. На рисунке дана схема процесса передачи в норадреналиновом синапсе. Сначала в три стадии происходит синтез норадреналина из аминокислоты тирозина, причем каждая стадия катализируется особым ферментом. Затем медиатор в сочетании с белками накапливается в пузырьках, примыкающих к мембране. Приходящий в аксонное окончание нервный импульс запускает приток ионов кальция, который вызывает высвобождение норадреналина из пузырьков в синаптическую щель. Молекулы медиатора связываются со специфическими рецепторными белками, включенными в постсинаптическую мембрану, запуская серию реакций, которая заканчивается кратковременными (электрическими) и долговременными воздействиями на воспринимающий нейрон. После этого действие норадреналина прекращается различными способами, в том числе быстрым возвращением медиатора в аксонное окончание и разрушением его ферментами. Выход некоторого количества норадреналина в синаптическую щель активирует пресинаптические рецепторы на аксонном окончании, вызывая выработку циклического АМФ, который активирует белковую киназу, стимулируя тем самым новую выработку норадреналина.
Известно около 30 разных веществ, относительно которых доказано или подозревается, что они играют роль медиаторов в головном мозгу, и каждое из них оказывает на нейроны характерный возбуждающий или тормозный эффект. Медиаторы распределены в мозгу не в случайном порядке, а локализованы в особых группах нейронов, аксоны которых идут к другим высокоспециализированным областям мозга. Наложение этих разнообразных химически закодированных систем на нейронные сети наделяет головной мозг еще одним измерением модуляции и специфичности.
За последние годы достигнуты значительные успехи в изучении различных медиаторных веществ (хотя многие из них, несомненно, еще не открыты), в составлении карт их распределения по мозгу и в выяснении молекулярных процессов синаптической передачи. Такими исследованиями установлено, что действие многих лекарственных веществ и нейротоксинов на поведение основано на их способности прерывать или модифицировать химическую передачу от нейрона к нейрону. В них есть также указания на то, что причинами психических болезней, возможно, окажутся в конечном счете нарушения функции специфических медиаторных систем мозга.
Что касается общего энергетического обмена, то из всех органов тела головной мозг является самым активным потребителем энергии, что отражается в его обильном кровоснабжении и интенсивном потреблении кислорода. Мозг настолько интенсивно использует кислород (50 миллилитров в минуту), что, составляя всего 2% общего веса тела, поглощает примерно 20% поступающего в организм кислорода.Такое огромное потребление энергии, как полагают, объясняется необходимостью поддерживать ионные градиенты по обе стороны нейронной мембраны, от чего зависит проведение импульсов в миллиардах нейронов мозга. Кроме того, это потребление энергии идет непрерывно: интенсивность метаболизма в мозгу относительно постоянна днем и ночью и иногда даже несколько возрастает во время фазы сна со сновидениями. Однако, чтобы не создалось ошибочного представления, следует сказать, что весь энергетический эквивалент метаболизма мозга составляет всего около 20 ватт.
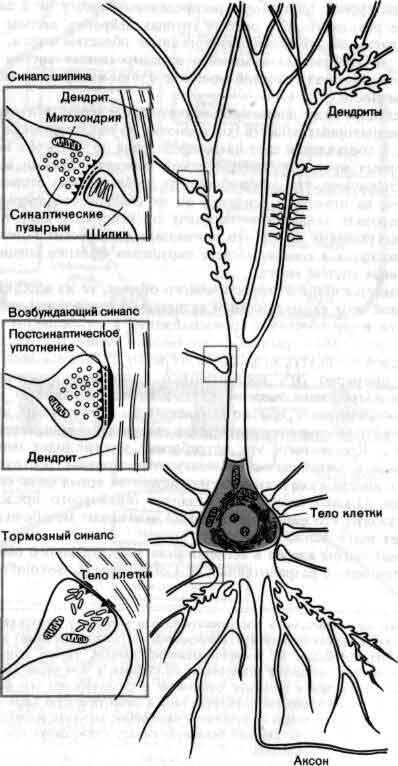
Синапсы на типичном нейроне в головном мозгу являются либо возбуждающими, либо тормозными, в зависимости от типа выделяющегося в них медиатора. Они различаются морфологически под электронным микроскопом: для возбуждающих синапсов характерны сферические пузырьки и сплошное утолщение постсинаптической мембраны, а для тормозных — уплощенные пузырьки и несплошное утолщение мембраны. Синапсы можно также классифицировать по их положению на поверхности воспринимающего нейрона — на теле клетки, на стволе или «шипике»дендрита, или на аксоне.
Важным шагом вперед в исследовании энергетического обмена мозга является метод, разработанный Л. Соколовым (L. Sokoloff) с сотрудниками в Национальном институте охраны психического здоровья. Этот метод позволяет визуально определять интенсивность энергетического обмена в клетках мозга. Нейроны приспосабливают потребление глюкозы к удовлетворению своих метаболических потребностей в данный момент. Следовательно, в активном состоянии они поглощают ее быстрее, чем в покое. Поглощенная глюкоза обычно претерпевает быстрые превращения; ее химический аналог, 2-дезоксиглюкоза, поглощается клетками точно так же, но не подвергается метаболизму. Если ввести в кровь дезоксиглюкозу с радиоактивной меткой, то она накопится в нейронах, и скорость накопления послужит показателем метаболической активности клетки. Накопление радиоактивной дезоксиглюкозы можно установить и измерить, накладывая тонкие срезы замороженного мозга на радиочувствительную пленку. На проявленной пленке выявляются участки, богатые меченым веществом. Эта методика открыла совершенно новую область исследований мозга, поскольку она позволяет установить, какие клетки в головном мозгу были активны во время данного эксперимента. Например, воздействуя световым стимулом (вспышка) на правый или левый глаз, можно определить, какие именно области мозга получают зрительную афферентацию от того или другого глаза.
В отличие от других органов тела, способных использовать разные виды «топлива» (сахара, жиры и аминокислоты), нейроны используют только глюкозу крови. Кроме того, в отличие от таких тканей, как мышцы, способных кратковременно функционировать в отсутствие кислорода, головной мозг полностью зависит от окислительного метаболизма. Если приток окисленной крови к мозгу прекратится, то через 10 секунд наступит потеря сознания, а затем появятся стойкие нарушения. Подобный же эффект вызывает любое состояние, сопровождающееся понижением содержания глюкозы в крови, например гипогликемия у больного диабетом, вызванная передозировкой инсулина. Хотя тонкие регуляционные механизмы обеспечивают постоянство кровяного давления и постоянный уровень кислорода и глюкозы в крови, очевидно, что чрезвычайная гибкость поведения, ставшая возможной благодаря большим размерам и емкости головного мозга млекопитающих, приобретена в процессе эволюции ценой высоких метаболических затрат.
Что касается клеток, то нейроны чрезвычайно чувствительны: их активность нарушается токсическими веществами, попавшими в кровоток, а также мелкими молекулами, обычно присутствующими в крови, например аминокислотами. Такая чувствительность, возможно, служит причиной того, почему головной мозг отделен от общего кровообращения избирательной фильтрационной системой, называемой гематоэнцефалическим барьером. Эффективность этого барьера объясняется относительной непроницаемостью кровеносных сосудов головного мозга и наличием плотного слоя глиальных клеток (опорных мозговых клеток) вокруг них. Хотя такие мелкие молекулы, как молекулы кислорода, легко проникают сквозь барьер, большинство более крупных молекул, необходимых клеткам мозга, например молекулы глюкозы, должны захватываться активно с помощью специальных транспортных механизмов. О гематоэнцефалическом барьере следует помнить при создании лекарственных средств, действующих непосредственно на мозг: для того чтобы такие вещества проходили через барьер, они должны состоять из очень малых молекул или быть легко растворимыми в жировых мембранах глиальных клеток. Несколько участков мозга не защищены тематоэнцефалическим барьером; к ним относятся такие структуры, которые специфически реактивны в отношении содержащихся в крови гормонов, и такие, функция которых состоит в регуляции химического состава крови.
Внутри отдельных нейронов существуют свои транспортные проблемы, связанные с тем, что клетка частично состоит из длинных, тонких волокон. Длина аксона, проводящего нервный импульс от тела нейрона, может исчисляться в миллиметрах или сантиметрах. Нейроны в мозгу взрослого животного незаменяемы, сохраняются на протяжении всей жизни; поэтому должны существовать механизмы возобновления всех их компонентов. Для выполнения этого требования клетка должна синтезировать ферменты и другие сложные молекулы, а такой синтез возможен только в районе клеточного ядра, т. е. в теле нейрона. Поэтому для возобновления своих компонентов аксону необходимо средство транспортировки их внутри клетки на значительные расстояния. И действительно, от тела клетки по всей длине аксона непрерывно движутся белки и другие составные части.
Феномен аксонного транспорта был открыт более 30 лет назад П. Вейсом (P. Weiss) и его сотрудниками из Чикагского университета. До этого все считали, что аксоплазма - желеобразная жидкость внутри аксона - служит лишь неподвижной механической опорой для возбудимой мембраны, по которой распространяется нервный импульс. Но когда Вейс пережал аксон в определенной точке, он отметил, что через несколько дней волокно разбухло около места сжатия со стороны тела клетки и сузилось с противоположной стороны. Когда же он устранил сжатие, задержанная аксоплазма возобновила свое течение.
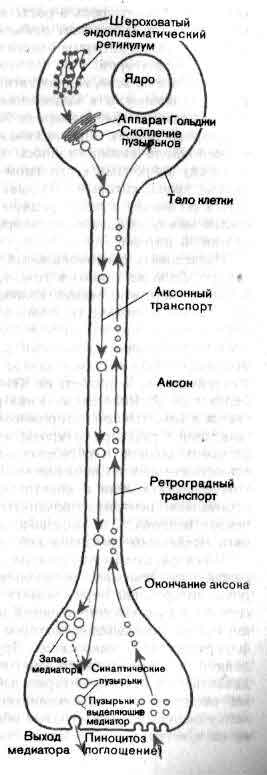
Аксонный транспорт обеспечивает передвижение компонентов клетки, таких, как пузырьки и ферменты, от места их образования в теле нейрона к окончанию аксона на расстояние в несколько миллиметров или сантиметров. В обратном направлении - от окончаний аксона к телу клетки-переносятся вещества, нужные для поддержания нормального состояния нейрона. Аксонный транспорт можно использовать, чтобы проследить ход проводящих путей, наблюдая за движением вдоль аксонов меченных изотопами молекул или таких ферментов, как пероксидаза хрена.
Известно, что аксоплазма служит артерией для оживленного движения молекул в обе стороны между телом нервной клетки и окончаниями ее аксона. В этом принимают участие несколько разных систем, в том числе система медленного транспорта, когда вещество течет от тела клетки со скоростью около одного миллиметра в сутки, и система быстрого транспорта, когда вещество движется в обе стороны со скоростью от 10 до 20 сантиметров в сутки. Система медленного транспорта - это течение основной массы аксоплазмы с переносом компонентов, играющих важную роль в росте и регенерации аксона; система быстрого транспорта охватывает движение более специализированных компонентов клетки, к которым относится ряд ферментов, участвующих в выработке медиаторов.
Пока еще не ясно, как достигаются эти разные скорости транспорта, но, по-видимому, и в медленном и в быстром механизмах участвуют многочисленные фибриллярные белки аксона, видимые на электронных микрофотографиях. Прослеживая транспорт белков с радиоактивной меткой вдоль аксона, удалось точно установить анатомические связи между нейронами в головном мозгу. При помощи фермента пероксидазы хрена, который обладает особым свойством быстро передвигаться по аксону в ретроградном направлении, можно составить карту связей между окончанияминейрона в одном участке мозга и телом клетки — в другом.
Исследовать функциональный химизм головного мозга чрезвычайно трудно. Дело не только в том, что медиаторные вещества содержатся в нем в ничтожно малых количествах, но и в том, что ткань мозга структурно и химически очень сложна, и выделить для исследования определенную медиаторную систему не легко. Однако для преодоления этих устрашающих препятствий разработаны новые методики. Большой шаг вперед был сделан в начале 1960-х годов благодаря открытию В. Уиттейкера (V. Whittaker) из Кембриджского университета и Э. де Робертиса (Е. de Robertis) из Университета в Буэнос-Айресе. Оно заключается в том, что при осторожном разрушении ткани мозга путем гомогенизации в растворе сахарозы многие нервные окончания отрываются от своих аксонов и образуют особые замкнутые частицы, названные «синаптосомами». Синаптосомы содержат механизмы синтеза, хранения, высвобождения и инактивации медиатора, связанные с нервным окончанием; центрифугированием их можно очистить от других компонентов нейрона. Эта методика дала нейрохимикам возможность изучать механизмы синаптической передачи в пробирке.
Пожалуй, самым плодотворным методическим достижением явилась разработка приемов, позволяющих избирательно окрашивать нейроны, содержащие определенный медиатор. Один из подходов к этому состоит в том, что естественный медиатор преобразуется в его флуоресцирующее производное, которое светится в ультрафиолетовых лучах флуоресцентного микроскопа. Другой подход состоит в том, что в головной мозг подопытного животного вводят молекулы медиатора с радиоактивной меткой, которые избирательно захватываются нейронными окончаниями, выделяющими этот медиатор; такие содержащие метку окончания можно затем обнаружить, поместив тонкие срезы ткани на чувствительную пленку. Третий подход использует высокую специфичность антител. Фермент, участвующий в синтезе данного медиа-гора, выделяют из ткани мозга и вводят в организм животного, где он вызывает образование антител, способных специфически реагировать с данным ферментом. Затем антитела подвергают очистке, метят флуоресцирующим красителем или используют какую-либо другую метку и применяют для избирательного окрашивания нейронов, содержащих данный фермент.
Эти методики избирательного окрашивания позволили получить поток сведений о детальном анатомическом распределении отдельных медиаторов в сложных нейронных цепях головного мозга. Они показали, что медиаторы распределены не диффузно по всей ткани мозга, а в высшей степени локально в ограниченных центрах и путях. Из всех медиаторов наиболее полно составлены карты для моноаминов - норадреналина, дофамина и серотонина (названных так. потому что все они содержат по одной аминогруппе). Б. Фальк (В. Falck) из Лундского университета и Н. Хжлларп (N. Hillarp) из Каролинского института в Швеции впервые показали, что воздействуя формальдегидом или глиоксиловой кислотой, можно превратить эти медиаторы в их флуоресцирующие производные, и тогда нейроны, содержащие моноамины, дают зеленую или желтую флуоресценцию. Такие работы показали, что многие содержащие норадреналин клетки мозга сосредоточены в стволе и образуют скопление, известное как locus coeruleus. Аксоны этих нейронов сильно ветвятся и проецируются в различные области — гипоталамус, мозжечок и передний мозг. Норадреналиновые нейроны оказались причастными к поддержанию бодрствования, к системе поощрения (центр удовольствия), к сновидениям и к регуляции настроения.
Нейроны, содержащие моноамин дофамин, сосредоточены в областях среднего мозга, известных под названиями substantia nigra и вентральной покрышки. Многие содержащие дофамин нейроны посылают свои аксоны в передний мозг, где, как предполагают, участвуют в регуляции эмоциональных реакций. Другие дофаминовые волокна оканчиваются в головном мозгу, в области полосатого тела. Здесь дофамин, очевидно, играет решающую роль в регуляции сложных движений. Дегенерация приходящих в эту область дофаминовых волокон приводит к ригидности мышц и тремору, столь характерным для болезни Паркинсона.
Моноаминовый медиатор серотонин сосредоточен в области ствола мозга, где находятся так называемые «ядра шва». Нейроны этого центра проецируются в гипоталамус, таламус и многие другие области мозга. Как полагают, серотонин участвует в терморегуляции, сенсорном восприятии и засыпании.
Идентифицировано еще много других медиаторов, часть из них считаются «предполагаемыми», поскольку их причастность к синаптической передаче в мозгу еще не доказана. Так, например, некоторые аминокислоты - блоки, из которых построены белки, - по-видимому, действуют как медиаторы. Обычно содержащиеся в организме в большом количестве аминокислоты - глутаминовая и аспарагиновая - оказывают сильное возбуждающее действие на большинство нейронов, и весьма возможно, что они являются наиболее распространенными возбудительными медиаторами в синапсах головного мозга. Известно, что самая простая из всех аминокислот, глицин, служит тормозным медиатором в спинном мозгу.




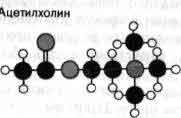



Медиаторы представляют собой вещества, молекулы которых имеют небольшой размер и положительно заряженный атом азота. Каждое такое вещество оказывает характерное либо возбуждающее, либо тормозное действие на нейроны; правда, есть медиаторы, которые действуют как возбуждающие в одной области мозга и как тормозные в другой. Принадлежность гистамина и таурина к медиаторам точно не доказана. Согласно принципу Дэйла, на всех окончаниях одного аксона выделяется один и тот же медиатор. Однако недавно обнаружены исключения из этого правила.
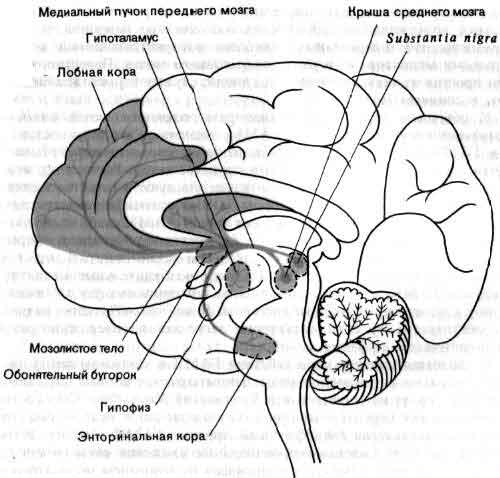
Дофаминовые проводящие пути показаны на рисунке схематически. Тела нейронов, содержащих дофамин, образуют скопления в двух небольших участках среднего мозга - substantia nigra и покрышке. От этих нейронов отходят широко ветвящиеся волокна, которые оканчиваются в полосатом теле, регулирующем двигательную активность, и в лимбической системе переднего мозга, связанной с эмоциями. Небольшая группа дофаминовых нейронов в гипоталамусе регулирует также секрецию гормонов гипофизом. Дофамин связывают с двумя заболеваниями: дефицит медиатора в полосатом теле создает ригидность и тремор, характерные для болезни Паркинсона, а избыток дофамина в лимбической системе переднего мозга, возможно, причастен к шизофрении.
К обычным тормозным медиаторам головного мозга относится гамма-аминомасляная кислота (ГАМК), которая не входит в состав белков. ГАМК занимает среди кислот особое место — она вырабатывается почти исключительно в головном и спинном мозгу. Вычислено, что не менее трети синапсов головного мозга используют в качестве медиатора ГАМК. Содержащие ее нейроны можно идентифицировать двумя способами: пометив их либо радиоактивной ГАМК, либо с помощью антител, специфичных в отношении глутаматдекарбоксилазы - фермента, который катализирует синтез ГАМК. Интересно отметить, что глутаминовая кислота считается кандидатом в возбудительные медиаторы, тогда как ГАМК, которая отличается от нее лишь на одну химическую группу, служит тормозным медиатором. Ясно, что небольшие различия в молекулярном строении медиаторов могут давать совершенно разные физиологические эффекты.
Исследование механизмов действия ГАМК в головном мозгу было стимулировано в последние годы открытием, сделанным недавно Т. Перри (Т. Perry) из Университета Британской Колумбии. Оно состоит в том, что для хореи Гентингтона - наследственного неврологического синдрома — характерен специфический дефицит ГАМК в мозгу. Возникающие при этой болезни непроизвольные движения связаны с наступающим в среднем возрасте постепенным разрушением полосатого тела. Посмертные исследования показали, что поражение мозга состоит в гибели тормозных нейронов, которые в норме содержат ГАМК; можно предположить, что в основе болезни лежит дефицит этого медиатора. К сожалению, попытка лечить больных заменой недостающей ГАМК пока неосуществима, поскольку еще не созданы аналоги ГАМК, способные проходить сквозь гематоэныефалический барьер.
Недавно было высказано предположение, что на ГАМК направлено действие таких транквилизаторов, как диазепам и другие производные бензодиазепина. Из всех психотропных препаратов врачи предпочитают прописывать бензодиазепины, но механизм их действия до сих пор неизвестен. Имеющиеся данные позволяют думать, что эти препараты повышают эффективность ГАМК на уровне ее рецепторов в головном мозгу. Хотя в мозгу идентифицированы специфические центры связывания диазепама, явно отличные от рецепторов ГАМК, оба типа рецепторов, по-видимому, взаимодействуют друг с другом. Заманчиво предположить, что в головном мозгу содержится некое еще не открытое вещество, которое в норме действует на рецепторы диазепама; возможно, это естественное соединение, вызывающее или ослабляющее состояние тревоги.
Нейрохимики не только выяснили молекулярную структуру и анатомическое распределение разных медиаторов, но и достигли больших успехов в понимании точной последовательности биохимических явлений, участвующих в синаптической передаче. Процесс химической передачи проходит через ряд этапов: синтез медиатора, его накопление, высвобождение, взаимодействие с рецептором и прекращение действия медиатора. Каждый из этих этапов детально охарактеризован, и найдены препараты, которые избирательно усиливают или блокируют конкретный этап. Эти исследования позволили проникнуть в механизм действия психотропных лекарственных средств, а также выявить связь некоторых нервных и психических болезней со специфическими нарушениями синаптических механизмов.