Первым этапом химической передачи является синтез молекул медиатора в нервных окончаниях. Каждый нейрон обычно обладает только таким биохимическим аппаратом, какой ему нужен для синтеза медиатора одного типа, который выделяется из всех окончаний его аксона. Молекулы медиатора не синтезируются de novo, а изготовляются путем переработки предшественника — обычно аминокислоты — в результате ряда ферментативных реакций.
Создание медиатора требует или одного этапа ферментативного катализа (как в случае ацетилхолина), или до трех этапов (как адреналина). При синтезе норадреналина исходным веществом служит аминокислота тирозин, которая поступает в нервное окончание из крови. Тирозин сначала превращается в промежуточное соединение L-ДОФА; затем второй фермент превращает L-ДОФА в дофамин («полноправный» медиатор); а третий фермент превращает дофамин в норадреналин.
После выработки молекул медиатора они хранятся в окончании аксона в маленьких связанных с мембраной мешочках, называемых синаптическими пузырьками. В одном окончании могут быть тысячи синаптических пузырьков, а каждый пузырек содержит от 10000 до 100000 молекул медиатора. Пузырьки защищают их от разрушения ферментами внутри окончания.
Приход нервного импульса в окончание аксона вызывает высвобождение множества молекул медиатора из окончания в синаптическую щель. Механизм такого выделения остается спорным: одни исследователи полагают, что синаптические пузырьки прямо сливаются с пресинаптической мембраной и выбрасывают свое содержимое в синаптическую щель; другие утверждают, что подвижное скопление молекул медиатора выходит через специальные каналы. Но в любом случае известно, что нервный импульс запускает выход медиатора, повышая проницаемость нервного окончания для ионов кальция, которые устремляются в него и активируют механизм высвобождения молекул.
Вышедшие молекулы медиатора быстро проходят через наполненную жидкостью щель между окончанием аксона и мембраной воспринимающего нейрона. Здесь они взаимодействуют со специфическими рецепторами постсинаптической мембраны. Рецепторы фактически представляют собой крупные белковые молекулы, погруженные в полужидкую матрицу клеточной мембраны: части их торчат над и под мембраной подобно айсбергам. Выходящий на поверхность участок рецепторного белка и молекула медиатора имеют одинаковые очертания; они соответствуют друг другу наподобие ключа и замка.
Взаимодействие медиатора с его рецептором меняет трехмерную форму рецепторного белка, инициируя этим определенную последовательность событий. Это взаимодействие может вызвать возбуждение или торможение нейрона, сокращение мышечной клетки, а также образование и выделение гормона клеткой железы. Во всех этих случаях рецептор переводит сообщение, закодированное в молекулярной структуре медиатора, в специфическую физиологическую реакцию. Одни реакции, например сокращение произвольной мышцы, происходят за долю секунды; другие, например секреция гормона, занимают минуты, а иногда часы.
Многие рецепторы медиаторов имеют два функциональных компонента: центр связывания молекулы медиатора и пору, пронизывающую мембрану, избирательно проницаемую для определенных ионов. Связываясь с рецептором, медиатор меняет его форму так, что пора открывается и ионы, находящиеся внутри и снаружи клеточной мембраны, перемещаются вдоль градиента концентрации, оказывая этим возбуждающий или тормозный эффект на частоту импульсации нейрона. Будет ли электрический потенциал, создаваемый медиатором, возбудительным или тормозным, зависит от того, какие именно ионы перемещаются, и от направления их движения. Ацетилхолин является возбуждающим медиатором в синапсе между нервом и мышцей, потому что он заставляет положительно заряженные ионы натрия входить в клетку и понижать ее отрицательный потенциал покоя. ГАМК, напротив, соответствует рецептор, у которого пора избирательно проницаема для отрицательно заряженных ионов хлора. Когда эти ионы входят через открытые поры в воспринимающую клетку, они повышают трансмембранный потенциал и на время инактивируют клетку.
Другие медиаторы, например дофамин и норадреналин, по-видимому, действуют посредством более тонкого механизма. В середине 50-х годов И. Сьюзерленд мл. (Е. Sutherland, Jr.) и его коллеги по Университету Кейса Западной резервной территории показали, что эти и другие медиаторы повышают или снижают концентрацию «второго посредника» в воспринимающих клетках. Затем второй посредник передает электрические или биохимические эффекты медиатора, или «первого посредника». Позднее в работе, которая в 1971 г. принесла Сьюзерленду Нобелевскую премию по физиологии и медицине, он установил, что второй посредник представляет собой небольшую молекулу циклического аденозинмонофосфата (цАМФ).
Согласно гипотезе Сьюзерленда, белковый рецептор норадреналина (и многих других медиаторов) соединяется в мембране клетки-мишени с ферментом аденилатциклазой, которая катализирует превращение в клетке богатой энергией молекулы аденозинтрифосфата (АТФ) в цАМФ. Аденилатциклаза обычно неактивна, но, когда норадреналин связывается с постсинаптическимрецептором, фермент автоматически включается и внутри клетки начинается быстрое превращение АТФ в цАМФ. Затем цАМФ действует на биохимический аппарат клетки, вызывая физиологическую реакцию, характерную для данного медиатора.
Система второго посредника сходна поэтому с эстафетой, в которой медиатор передает свое сообщение циклическому АМФ в мембране клетки. Разумеется, сигнал передается не одной, а многим тысячам молекул цАМФ, которые генерируются активированной аденилатциклазой, связанной с каждым занятым рецептором. В результате очень слабый сигнал, создаваемый взаимодействием медиатора с рецептором, усиливается внутри клетки в несколько тысяч раз благодаря интенсивному образованию цАМФ.
Приложение теории Сьюзерленда о втором посреднике к функции головного мозга является одной из самых увлекательных областей современной биохимии. В 1971 г. Ф. Блум (F. Bloom) с сотрудниками в Национальных институтах здравоохранения показали, что цАМФ способен влиять на генерацию сигналов в нейронах. Позднее П. Грингард (P. Greengard) и его группа в Медицинской школе Йельского университета пришли к заключению, что цАМФ участвует в синаптическом действии нескольких медиаторов, в том числе норадреналина, дофамина, серотонина и гистамина. Грингард предложил обобщающую гипотезу, согласно которой цАМФ активирует специфические ферменты в постсинаптической клетке, именуемые белковыми киназами; затем эти ферменты катализируют внедрение фосфатных групп в специальные белки в мембране нейрона, изменяя проницаемость мембраны для ионов и тем самым изменяя уровень возбудимости клетки-мишени. Поскольку система второго посредника работает сравнительно медленно по шкале времени нейронных событий, она больше всего пригодна для участия в более длительных эффектах медиаторов мозга, таких, как медленные сдвиги мембранного потенциала и, возможно, образовании следов долговременной памяти. Как только цАМФ передал свое сообщение дальше, он инактивируется в клетке под действием фермента фосфодиэстеразы. Поэтому препараты, ингибирующие этот фермент, повышают уровень цАМФ в постсинаптических клетках и усиливают действие медиатора.
Итак, по-видимому, существуют два основных типа медиаторных рецепторов: быстро действующие рецепторы, которые осуществляют передачу информации, регулируя проницаемость ионной поры, и медленно действующие рецепторы, которые вызывают образование второго посредника; последний в свою очередь опосредует эффекты, производимые медиатором в постсинаптическом нейроне. Для многих медиаторов имеется по два и более типов рецепторов. Например, реакция на ацетилхолин в синапсе между мотонейроном и мышечной клеткой осуществляется простым током ионов натрия через мембрану. Но в головном мозгу эффекты ацетилхолина в большинстве своем опосредуются, по-видимому, молекулой еще одного второго посредника, циклического гуанозинмонофосфата, или цГМФ. Точно так же недавно полученные данные позволяют думать, что дофамин действует на уровне двух разных типов рецепторов в головном мозгу: рецептора D1, который связан с системой второго посредника цАМФ, и рецептора D2, который с ней не связан.
Как только молекула медиатора свяжется со своим рецептором, она должна быть быстро инактивирована во избежание слишком длительного ее действия и нарушения точного контроля передачи. Нервные волокна способны проводить несколько сот импульсов в секунду только при условии, что постсинаптическая мембрана восстанавливает свой потенциал покоя за долю миллисекунды. Некоторые медиаторы инактивируются ферментами, находящимися в синаптической щели. Ацетилхолин, например, разрушается ферментом ацетилхолинэстеразой, которая за секунду расщепляет 25000 молекул медиатора. Норадреналин инактивируется в синапсе по совсем иному механизму.
Ю. Аксельрод (J. Axelrod) с сотрудниками из Национальных институтов здравоохранения установили, что выделившийся из аксонного окончания норадреналин снова быстро всасывается в окончание. Затем поглощенные молекулы норадреналина либо разрушаются ферментами - катехол-О-метилтрансферазой (КОМТ) и моноаминоксидазой (МАО), которые содержатся в нервном окончании, либо возвращаются обратно в синаптические пузырьки. Позднее такие же механизмы возврата были найдены и для других медиаторов — дофамина, серотонина и ГАМК. Возврат имеет то очевидное преимущество перед разрушением фермента, что молекулы медиатора сохраняются в течение нескольких циклов выделения и поглощения.
Выяснение этапов синаптической передачи пролило свет на способ действия психотропных препаратов. Некоторые из них действуют, либо усиливая, либо ослабляя высвобождение данного медиатора из аксонных окончаний. Например, под действием сильного стимулятора амфетамина в мозгу из нервных окончаний выделяется дофамин - медиатор, связанный с системами бодрствования и удовольствия. Чрезмерное применение амфетамина может привести к расстройствам мышления, галлюцинациям и мании преследования, т.е. к симптомам, очень сходным с теми, какие возникают при некоторых формах шизофрении. Эти и другие данные послужили основанием для предположения о том, что в основе симптомов шизофрении, возможно, лежит повышенная активность дофаминовых систем мозга.
Вызывает интерес также тот факт, что разнообразные лекарственные средства, созданные для лечения шизофрении, например хлорпромазин (торазин) и галоперидол (галдол), обладают общим свойством — они тесно связываются с дофаминовыми рецепторами головного мозга, тем самым мешая естественному медиатору активировать их. Это открытие оказалось одним из самых многообещающих в современном исследовании шизофрении. Последние данные позволяют думать, что шизофрения связана с избыточным образованием дофамина или с чрезмерной реактивностью в отношении медиатора в определенных участках головного мозга. Работами моей лаборатории в Отделе нейрохимической фармакологии Британского совета медицинских исследований и работами Т. Кроу (Т. Crow) в Центре клинических исследований Британского совета медицинских исследований в Лондоне, а также исследованиями Ф. Симена (Ph. Seeman) из Торонтского университета обнаружены необычно высокие концентрации дофамина в дофаминовых рецепторах в головном мозгу умерших больных шизофренией; это особенно относится к лимбической системе, т. е. тем областям мозга, которые ответственны за эмоциональное поведение. Поэтому возможно, что главной мишенью антипсихотических препаратов служат прежде всего пути дофамина в этих областях.
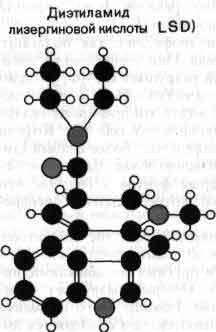
Многие психотропные препараты, возможно, действуют на уровне постсинаптических рецепторов, имитируя естественные медиаторы. Например, многие галлюциногены сходны по своей структуре с истинными медиаторами: мескалин похож на норадреналин и дофамин, a LSD и псилоцибин сходны с серотонином. Поэтому возможно, что перечисленные препараты действуют на моноаминовые механизмы, хотя способ их действия точно еще неизвестен. LSD выделяется своей необычайно высокой активностью: всего 75 микрограммов (едва видимой пылинки) достаточно чтобы вызвать галлюцинации.
Производные метилксантина кофеин и теофиллин, как полагают, действуют через систему второго посредника. Они специфически ингибируют фермент фосфодиэстеразу, который разрушает цАМФ, и таким образом в конечном счете увеличивают количество цАМФ, образующегося в ответ на действие медиатора. В результате эти препараты оказывают на головной мозг общее мягкое стимулирующее действие. Кофеин является главным действующим началом кофе и чая; более слабый стимулятор теофиллин содержится преимущественно в чае. Ежегодное потребление кофе и чая составляет миллиарды фунтов в год, так что метилксантины являются самыми широко потребляемыми препаратами.
Наконец, некоторые препараты потенцируют действие медиатора, блокируя его разрушение в синапсе. Одна из таких групп препаратов представлена ипрониазидом (марсилидом) и другими соединениями, ингибирующими фермент моноаминоксидазу, который разрушает норадреналин, дофамин и серотонин. Вследствие блокады этого фермента эффекты перечисленных моноаминов усиливаются, чем объясняется антидепрессивное действие таких лекарств. Вторая группа антидепрессантов - трициклические соединения - также усиливает действие норадреналина и серотонина в головном мозгу. Эти лекарственные средства, из которых лучше всего известны имипрамин (тофранил) и амитриптилин (элавил), блокируют обратный захват норадреналина и серотонина из синаптической щели; стимулятор кокаин, по-видимому, действует по тому же механизму. Указанные наблюдения говорят о том, что депрессия, по-видимому, связана с низким уровнем этих медиаторов.


Галлюциногены по своей структуре очень похожи на моноаминовые медиаторы; это позволяет допустить, что они оказывают сильные воздействия на сознание, имитируя действие этих естественных медиаторов на синаптические рецепторы в головном мозгу. Мескалин содержит в своей структуре бензольное кольцо подобно дофамину и норадреналину. а псилоцибин и LSD - индольное кольцо серотонина.
За последние годы, после того как в мозгу был обнаружен новый класс химических соединений нейропептиды, число известных систем химических посредников в головном мозгу резко возросло. Эти молекулы представляют собой цепочки из аминокислотных остатков (от двух до 39); они обнаружены в нейронах и предположительно являются медиаторами. Некоторые из них были вначале идентифицированы как гормоны, выделяемые гипофизом (АКТГ, вазопрессин), как локальные гормоны кишечника (гастрин, холецистокинин) или как гормоны, выделяемые гипоталамусом для регуляции выделения других гормонов гипофизом — люлиберина (гормона, стимулирующего выделение лютеинизирующего гормона), соматостатина.
Самыми новыми и вызывающими наибольший интерес нейропептидами являются энкефалины и эндорфины — вещества, встречающиеся в головном мозгу в норме и удивительно сходные с морфием — наркотиком, извлекаемым из снотворного мака. Вслед за открытием этих пептидов последовало осознание того факта, что некоторые области головного мозга с высокой степенью сродства связывают препараты группы опия. Рецепторы опия обнаружены путем определения связывания меченых его соединений с частицами нейронных мембран. Такие методики нанесения радиоактивных меток на рецепторы разработаны тремя группами исследователей, руководимыми С. Снайдером (S. Snyder) и К. Пертом (С. Pert) в Медицинской школе университета Джонса Гопкинса, Э. Саймоном (Е. Simon) из Медицинской школы Ньюйоркского университета и Л. Терениусом (L. Terenius) из Упсальского университета. Эти исследователи установили, что рецепторы опия сосредоточены в тех участках головного и спинного мозга млекопитающих, которые имеют отношение к восприятию и интеграции боли и эмоций.
Затем в 1975 г. Дж. Хьюз (J. Hughes) и X. Костерлиц (Н. Kosterlitz) в Абердинском университете выделили из головного мозга два естественных пептида, которые тесно связываются срецепторами опия, и назвали их энкефалинами. Оба энкефалина представляют собой цепочки, состоящие из пяти аминокислот в одинаковой последовательности, за исключением последней аминокислоты: в одном случае это метионин, а в другом — лейцин. Позднее из гипофиза были выделены другие морфиноподобные пептиды, названные эндорфинами. Недавно проведенные опыты показали, что некоторые процедуры, применяемые для устранения хронических болей,- иглоукалывание, прямая электрическая стимуляция головного мозга и даже гипноз, — возможно, действуют, вызывая выделение энкефалинов или эндорфинов в головном и спинном мозгу. Эта гипотеза основана на том, что эффективность всех этих процедур сильно снижается налоксаном (нарконом) — препаратом, который избирательно блокирует связывание морфина с рецептором опия.

Нейропептиды - короткие цепи аминокислот, обнаруженные в ткани мозга. Многие из них локализованы в аксонных окончаниях и высвобождаются из них под влиянием кальция, что дает основание считать их медиаторами. Однако нейропептиды отличаются от ранее идентифицированных медиаторов тем, что они организуют такие сложные явления, как жажда, память и половое поведение. Кроме того, они играют многообразную роль во многих других областях тела. Например, соматостатин подавляет выделение гормона роста гипофизом (у человека), регулирует секрецию инсулина и глюкагона поджелудочной железой и, по-видимому, функционирует в качестве медиатора в спинном и головном мозгу.
Показано, что многие нейропептиды, обнаруженные в головном мозгу млекопитающих, сосредоточены в окончаниях особых групп нейронов, и о некоторых из них известно, что выход их изаксонных окончаний зависит от кальция. Такие данные, а также наблюдение, что очень малые количества нейропептидов оказывают значительный эффект на активность нейронов или на поведение экспериментальных животных, в какой-то мере свидетельствуют в пользу того, что эти вещества вполне могут представлять собой новый класс медиаторов. Однако полученные данные еще не настолько убедительны, чтобы можно было считать это доказанным.
Пожалуй, самым первым кандидатом на статус медиатора среди нейропептидов является вещество Р - цепочка из 11 аминокислот. Оно присутствует во многих специфических нейронных путях в головном мозгу, а также в первичных сенсорных волокнах периферических нервов. Некоторые из этих сенсорных нейронов, тела которых лежат в сенсорных ганглиях по обе стороны спинного мозга, содержат вещество Р и выделяют его из своих аксонных окончаний в синапсах со спинальными нейронами. Поскольку вещество Р возбуждает те спинальные нейроны, которые легче всего реагируют на болевые стимулы, было высказано предположение, что оно служит сенсорным медиатором, специфически связанным с передачей информации о боли от периферических болевых рецепторов в центральную нервную систему.
Морфиноподобный пептид энкефалин тоже в изобилии содержится в мелких нейронах в той части спинного мозга, куда приходят волокна, содержащие вещество Р. Мы с Т. Джесселом (Т. Jessel) в Отделе нейрохимической фармакологии Британского совета медицинских исследований показали, что энкефалин и препараты опия способны подавлять выделение вещества Р из сенсорных волокон. Поэтому нейроны, содержащие энкефалин, могут регулировать поступление болевых сигналов в головной мозг, модулируя выделение вещества Р на уровне первого переключения в центральной нервной системе. Подобные же тормозные взаимодействия возможны и на более высоких уровнях мозга. Вещество Р - не единственный предполагаемый медиатор, локализующийся, как показано, в сенсорных нейронах; к таким же идентифицированным к настоящему времени веществам относятся ангиотензин, холецистокинин, соматостатин и глутаминовая кислота. Таким образом, по мере того как все больше становится известно о сенсорных медиаторах и их модуляционных механизмах в спинном мозгу, начинает возникать картина поразительной химической сложности.
Замечательное свойство нейропептидов мозга состоит в глобальном характере некоторых их эффектов. Введение очень малых количеств нейропептида (обычно прямо в головной мозг, чтобы обойти гематоэнцефалический барьер) вызывает у экспериментального животного сложное и весьма своеобразное поведение. Например, введение в мозг нескольких нанограммов нейропептида ангиотензина II приводит к тому, что животные интенсивно и долго пьют, хотя до этого не испытывали жажды. Другой пептид - гормон люлиберин, при введении его в головной мозг самке крысы создает характерное для самки половое поведение. Еще поразительнее, что, как показал Д. де Виед (D. de Wied) с сотрудниками из Утрехтского университета, введение малых количеств нейропептида вазопрессина лабораторным животным заметно улучшает запоминание действий, которым их обучили. Теперь проводятся предварительные клинические испытания этого препарата, чтобы установить, может ли он дать эффект в случаях потери памяти.

Гипотетический воротный механизм в первом синаптическом переключении, возможно, регулирует передачу информации о боли от периферических болевых рецепторов к головному мозгу. В задних рогах спинного мозга вставочные нейроны, содержащие пептидный медиатор энкефалин, образуют синапсы на аксонных окончаниях болевых нейронов, которые в качестве медиатора используют вещество Р. Выделяемый вставочными нейронами энкефалин тормозит выход этого вещества, из-за чего воспринимающий нейрон в спинном мозгу получает меньше возбуждающей стимуляции и поэтому посылает в головной мозг меньше связанных с болью импульсов. Такие опийные препараты, как морфий, по-видимому, связываются с незанятыми рецепторами энкефалина, имитируя подавление боли, производимое энкефалиновой системой.
Поэтому можно думать, что нейропептиды являются химическими посредниками, отличными от ранее идентифицированных медиаторов: по-видимому, они служат глобальным средством химического кодирования тех форм активности головного мозга, которые связаны с отдельными функциями поддержанием водно-солевого баланса, половым поведением, ощущениями боли или удовольствия. Неожиданным оказалось наблюдение, что биологически активные пептиды, первоначально обнаруженные в желудочно-кишечном тракте - гастрин, вещество Р, вазоактивный кишечный полипептид и холецистокинин - содержатся также и в центральной нервной системе. И наоборот, некоторые пептиды, сначала найденные в головном мозгу, позднее были обнаружены в кишечнике (соматостатин, нейротензин, энкефалины). Следовательно, можно думать, что эти пептиды выполняют множество ролей, действуя как локальные гормоны или медиаторы в желудочно-кишечном тракте и как глобальные медиаторы в головном мозгу. Р. Гиймен (R. Guillemin) из Института Солка высказал предположение, что разнообразные функции нейропептидов объясняются «оппортунизмом» эволюционного процесса, во время которого молекулу, обслуживающую одну функцию, в другое время и в другом месте можно приспособить для другой функции.

Локализация вещества Р в спинном мозгу обезьяны установлена путем обработки ткани специфическими антителами, меченными темным красителем. Краситель обнаруживается только в задних рогах спинного мозга, к которым подходят периферические болевые волокна. Микрофотография получена С. Хаитом (S. Hunt) из Кембриджского университета.
Ряд других веществ, по-видимому, играет роль модуляторов в межнейронных коммуникациях. На высоких уровнях головного мозга в его ткани найдены простагландины, которые состоят из пятичленного углеродного кольца и двух связанных с ним длинных углеродных цепей; простагландины оказывают разнообразное возбуждающее и тормозное действие на нейроны в зависимости от данной молекулярной структуры простагландина и характера клетки-мишени. В отличие от медиаторов с их быстрыми и кратковременными эффектами простагландины вызывают продолжительные сдвиги в поляризации нейронной мембраны; это говорит скорее об их модулирующей, а не о медиаторной функции. Возможно, что они действуют сообща с медиаторами, эффекты которых они мягко видоизменяют.
Еще одна группа веществ выполняет питательную функцию, а не функцию посредников. Считается, что эти «трофические» вещества выделяются нервными окончаниями и поддерживают жизнеспособность клетки-мишени; другие же трофические вещества захватываются нервными окончаниями, переносятся по аксону в ретрогралном направлении и питают этот же нейрон. Хорошо известный факт атрофии мышцы мосле нарушения ее иннервации, возможно, объясняется тем, что мышечные клетки перестают получать необходимые им трофические вещества. Некоторые дегенеративные заболевания мозга, возможно, возникают от того, что центральные нейроны перестают обмениваться трофическими веществами. В настоящее время из трофических веществ лучше всего описан фактор роста нервов — белок, который необходим для дифференцировки и выживания периферических сенсорных и симпатических нейронов и который, возможно, благоприятствует также поддержанию жизнеспособности центральных моноаминоэргических нейронов.
Наряду со все возрастающим числом химических медиаторов, становится также очевидным многообразие разных механизмов, посредством которых медиаторы производят свои эффекты. Например, вместо того чтобы прямо возбуждать или тормозить клетку-мишень, медиатор, выделяющийся из одного нервного окончания, может воздействовать на соседнее нервное окончание, усиливая или снижая выход медиатора из него. Ясно также, что для данного медиаторного вещества возможны Несколько типов рецепторов (на одни из них действуют, а на другие не действуют системы вторых посредников), чем объясняются неодинаковые возбудительные или тормозные эффекты данного медиатора в разных частях мозга. Даже утвердившееся представление [впервые высказанное Г. Дэйлом (Н. Dale)] о том, что из всех своих окончаний нейрон выделяет один и тот же химический медиатор, может утратить свою нерушимость: установлено, что с норадреналином или серотонином в том же нейроне сосуществует ряд нейропептидов. Функциональное значение таких двойных медиаторных систем пока еще не раскрыто. Кроме того, остаются во многом неизвестными те конкретные химические нарушения, которые лежат в основе таких обычных заболеваний, как эпилепсия, старческое слабоумие, алкоголизм, шизофрения и депрессия. Хотя исследование медиаторных систем мозга уже очень много раскрыло в химических механизмах обучения, памяти, сна и настроения, тем не менее ясно, что самые волнующие открытия еще впереди.
Д. ХЬЮБЕЛ, Т. ВИЗЕЛЬ
Центральные механизмы зрения
Путем изучения активности и пространственной организации нейронов первичной зрительной коры выявляется функциональная схема, которая может лежать в основе переработки сенсорной информации в коре
Рассматриваемая как продукт эволюции кора большого мозга должна считаться одним из самых больших достижений в истории всего живого. У позвоночных, стоящих ниже млекопитающих, кора мозга едва выражена, если вообще можно говорить о ее существовании. Внезапно приобретя внушительные размеры у низших млекопитающих, она начинает доминировать в мозгу хищников и взрывоподобным образом увеличивается у приматов; у человека она почти полностью охватывает остальную часть мозга, имея тенденцию затмевать другие отделы. Степень зависимости животного от какого-то органа является показателем важности последнего, причем показатель этот даже более убедителен, чем размер, а зависимость от коры быстро нарастала по мере эволюции млекопитающих. Мышь без коры ведет себя относительно нормально, по крайней мере при поверхностном исследовании; в то же время человек без коры - это почти растение, бессловесное, невидящее, бесчувственное.

Колонки глазодоминантности, одна из двух основных систем, характеризующих функциональную организацию первичной зрительной коры, видны на этом радиоавтографе как периодические светлые участки (срез коры обезьяны-макака, рассматриваемый в темном поле). Данные колонки (в действительности, искривленные подслои коры, видимые здесь в поперечном сечении на срезе мозга, перпендикулярном поверхности) - это участки, в которых все нейроны реагируют более активно на правый глаз, чем на левый; темные участки, разделяющие светлые области, - колонки предпочтения левого глаза. Для получения этого радиоавтографа в правый глаз анестезированного животного была введена меченая аминокислота. Поглощенная телами клеток сетчатки аминокислота доставлялась к клеткам хоры через коленчатое тело - промежуточную станцию мозга. Срез коры был покрыт фотографической эмульсией, которая экспонировалась в течение нескольких месяцев, а затем была проявлена. Засвеченные зерна серебра над радиоактивными участками образовали рассеивающие свет области, которые соответствуют колонкам глазодоминантности.

Колонки глазодоминантности, видимые в фас на радиоавтографе, полученном методом аксонного транспорта, для среза, параллельного, а не перпендикулярного поверхности первичной зрительной коры. Как можно видеть на предыдущем радиоавтографе, самые яркие метки находятся в одном слое складчатой коры - слое IV. Это уровень, на котором оканчиваются аксоны, несущие информацию в кору; соответственно здесь и накапливается метка. Данный срез был сделан в плоскости, тангенциальной колоколовидной поверхности коры, сразу под слоем IV, который в связи с этим выглядит как кольцо с текстурой из приблизительно параллельных ярких полос. Это выявленные радиоактивными метками области глазодоминантности, которые видны теперь сверху, а не сбоку. Действительная ширина областей глазодоминантности, как правило, составляет около 0,4 миллиметра.