Как головной и спинной мозг управляют движениями тела? Мозг не только посылает команды мышцам, но и получает по обратной связи сигналы, которые помогают ему согласовывать эти команды
Одно из первых сведений, полученных более ста лет назад, об управлении движением со стороны головного мозга состояло в том, что движения тела могут быть вызваны сигналами, приходящими в спинной мозг из специальной области головного мозга - моторной зоны коры больших полушарий. Движения имеют широкий диапазон - от мышечных координации, требуемых для грубой ручной работы или быстрого перемещения всего тела, до тонких движений пальцев при хирургических операциях, выполняемых под микроскопом.

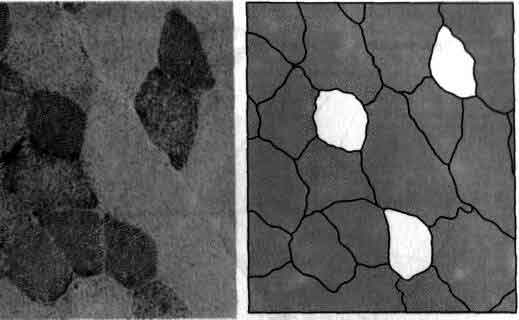
Три белые «тени» на микрофотографии представляют собой метки, специально созданные, чтобы облегчить исследование важного аспекта связи между головным мозгом и движением, а именно химизма мышечного сокращения, следующего за импульсацией мотонейрона. На микрофотографии показан поперечный срез одной из мышц конечности кошки. «Тени» образованы отдельными мышечными волокнами в одной двигательной единице. Предварительное изучение этой единицы показало, что она относится к «медленной» мышце, т. е. такого рода мышце, которая развивает небольшую силу, но функционирует не утомляясь. Эти три волокна превратились в метки в результате длительной стимуляции мотонейрона, который управляет их сокращениями, что привело к истощению запаса гликогена - особой формы хранения глюкозы, которая служит источником энергии для работы мышцы. При окрашивании среза все мышечные волокна с нормальным содержанием гликогена стали розовыми. На следующих рисунках показаны другие срезы той же мышцы с теми же тремя метками. Они были окрашены, чтобы определить связь между химическими и механическими свойствами мышечных волокон. Микрофотографии получены Р. Берком (R. Burke) и П. Церисом (P. Tsairis) в Национальных институтах здравоохранения.
Эта выходная активность моторной коры сама является результатом сигналов, поступающих из других пунктов - не только от других областей коры, например тактильной, но и от подкорковых структур мозжечка и базальных ганглиев, которые посылают сигналы в моторную кору еще через одно подкорковое образование — таламус. Основная часть современных исследований мозговых механизмов движения направлена на лучшее понимание того, как сигналы, приходящие от различных корковых и подкорковых структур, объединяются в контроле над конечными выходами из моторной коры к спинному мозгу и оттуда к мышцам. В настоящей статье будет рассмотрен современный уровень наших знаний, которые имеют важное значение по двум причинам. Во-первых, они связаны с фундаментальными проблемами общей организации головного мозга. Во-вторых, они имеют отношение к лечению и, возможно, к предупреждению таких неврологических заболеваний, как болезнь Паркинсона и хорея Гентингтона (две болезни из числа тех, при которых затронуты базальные ганглии), различные проявления инсульта, рассеянного склероза, а также многих других нарушений, возникающих при повреждении мозжечка.
Каковы элементарные условия для выполнения движения? Первое — это мышца, второе — это сигнализирующая система, которая вызывает упорядоченное сокращение мышцы. Если начать с мышц, то надо сказать, что не все они работают одинаково. Рассмотрим мышцы глаза и руки у человека. Глазные мышцы должны работать с высокой скоростью и большой точностью, быстро ориентируя глазное яблоко в пределах нескольких дуговых минут. В то же время глазной мышце не приходится справляться с такими внешними задачами, как поднимание груза. Тонкое управление, требуемое при движении глаза, требует высокого иннервационного индекса — отношения числа нейронов, аксоны которых оканчиваются на наружной мембране мышечных клеток, к числу мышечных клеток.
Для глазной мышцы иннервационный индекс составляет 1:3; это значит, что аксонные окончания одного мотонейрона выделяют свой медиатор не более чем на три отдельные мышечные клетки. (Мотонейрон — это такой нейрон, тело которого лежит в спинном мозгу, а аксон оканчивается на мембране мышечной клетки.) По-иному обстоит дело с мышцами руки: аксонные окончания одного мотонейрона, например иннервирующего бицепс, могут действовать своим медиатором на сотни мышечных волокон, и поэтому у такой мышцы иннервационный индекс составляет всего 1:100. В результате действие одной двигательной единицы мышцы конечности — одно быстрое сокращение (twitch), возникающее под влиянием одного импульса, вызывающего выделение медиатора из окончаний одного мотонейрона, — соответственно оказывается грубым.
Двигательные единицы мышц различаются также по тому, насколько они подвержены утомлению. На одном конце шкалы лежат двигательные единицы медленного сокращения, способные длительно функционировать без утомления. Такие единицы могут быть активными в течение длительного времени, но развивают сравнительно небольшое мышечное напряжение. На другом конце шкалы находятся двигательные единицы быстрого сокращения; они могут создавать высокие пики мышечного напряжения, но быстро утомляются. Такие единицы обычно иннервируются мотонейронами с диаметром аксонов и скоростью проведения нервного импульса выше средних.
В одной и той же мышце содержатся волокна и быстрых и медленных двигательных единиц. В 1968 г. шведские исследователи Э. Кугельберг (Е. Kugelberg) и Л. Эдстром (L. Edstrom) нашли способ определять, какие отдельные моторные волокна относятся к данной двигательной единице. Длительной стимуляцией аксона одного мотонейрона эти исследователи вызывали продолжительное сокращение мышечных волокон одной двигательной единицы. Сокращение приводило к истощению в отдельных мышечных волокнах запаса гликогена, который является источником энергии. При специальном окрашивании ткани на гликоген волокна истощенной двигательной единицы принимали вид белых «теней», рассеянных среди розовых волокон с нормальным запасом гликогена.
Такой гистохимический эффект представляет собой демонстрацию биохимической реакции в живой микроанатомической структуре. Так, применив подход Кугельберга и Эдстрома, Р. Берк (R. Burke) с сотрудниками из Национальных институтов здравоохранения показал с помощью гистохимического окрашивания, что «быстрые» мышечные единицы, используя в качестве источникаэнергии аденозинтрифосфат (АТФ), расщепляют его ферментативным путем быстрее, чем это происходит в «медленных» двигательных единицах. Это ферментативное расщепление считается одним из важных факторов, определяющих присущую мышце скорость сокращения. Таким образом, гистохимические данные помогают объяснить различия в скорости сокращения. Равным образом, гистохимическое исследование других ферментов — тех, которые расщепляют сахара и жиры, — помогает объяснить весьма существенные различия в утомляемости между двумя видами двигательных единиц.
Каково значение этих противоположных свойств двигательных единиц для организации движения? Посмотрим, как двигательные единицы мышцы последовательно «вовлекаются» в процесс движения. В общем мышечное напряжение регулируется двумя путями. Первый состоит в контроле над числом двигательных единиц, вовлекаемых в активность. Второй — в регуляции частоты импульсации вовлеченных единиц. Первыми вовлекаются единицы медленного сокращения, не склонные к утомлению и развивающие сравнительно слабое напряжение. Последними вовлекаются двигательные единицы быстрого сокращения, т. е. те, которые дают высокие пики напряжения, но быстро утомляются.

Второй срез той же мышцы был окрашен для определения относительной способности мышечных белков расщеплять аденозинтрифосфат (АТФ). Темная окраска свидетельствует о более высокой расщепляющей активности. Три волокна-метки очень слабо окрашены (схема справа); такая низкая активность характерна для медленных мышечных волокон.

Третий срез окрашен, чтобы показать способность мышечных белков расщеплять АТФ после их предварительной обработки кислотой. Обратное соотношение интенсивности окраски по сравнению со вторым срезом дает дальнейшие сведения о химизме мышечного волокна.
Четвертый срез был окрашен, чтобы показать относительную способность мышечных волокон к окислительному метаболизму, определяемому по наличию ключевого фермента в митохондриях клетки. Три волокна медленной двигательной единицы (см. схему) находятся среди интенсивно окрашенных волокон; картина согласуется с представлением о меньшей утомляемости таких двигательных единиц.
Э. Хеннемен (Е. Henneman) из Гарвардской медицинской школы внес важный вклад в общее понимание порядка вовлечения в активность отдельных двигательных единиц. Он отметил, что напряжение мелких мышц создается и точно контролируется избирательной мобилизацией разного числа мелких двигательных единиц. Между мелкими и более крупными единицами существует немалая разница. Например, самая крупная двигательная единица в икроножной мышце человека развивает напряжение в 200 раз больше, чем самая мелкая. Когда требуется общее усиление работы мышц, то, как установил Хеннемен, в активность вовлекаются более крупные двигательные единицы, что создает большие элементарные приросты напряжения. Это означает, что по мере увеличения общего напряжения оно создается меньшим числом дополнительных единиц. Разумеется, когда обстоятельства требуют резкого повышения общего напряжения мышцы, двигательные единицы вовлекаются не последовательно, а активируются практически одновременно.
Так обстоит дело с разными видами мышц и их двигательными единицами. Теперь посмотрим, что заставляет эти единицы сокращаться. Мышечные сокращения совершаются благодаря тому, что медиатор ацетилхолин, который выделяется в нервно-мышечном соединении при каждом импульсе, исходящем от мотонейрона, способен вызвать импульс в мышечной клетке. Блокирование передачи на уровне соединения, например при помощи алкалоида кураре, препятствует мышечному сокращению. Такую блокаду воспроизводили и у людей. Испытуемым вводили кураре в условиях искусственного дыхания. Во время паралича, вызванного кураре, выделение ацетилхолина аксонами мотонейронов продолжается, но блокируется взаимодействие медиатора с соответствующими рецепторами на мембране мышечной клетки; в результате мышцы перестают реагировать на команды, поступающие от коры больших полушарий. Испытуемые продолжали мыслить и чувствовать, но внешние проявления деятельности мозга исчезали. Речь, выражение лица, способность направлять взор — все эти формы поведения зависят от сокращения мышц.

Головной мозг макака, вид сверху. Помечены разные участки моторной коры больших полушарий. Окрашенная полоса обозначает ту часть мозга, которая удалена, чтобы показать подкорковые структуры на соседнем рисунке.
Многие современные представления о механизмах движения возникли на основе трудов английского физиолога Шеррингтона, который в начале века занимался изучением функции мотонейронов в некоторых рефлекторных формах двигательной активности, таких, как чесание и ходьба. Сигналы, идущие от многих различных областей головного мозга, часто воздействуют на несколько одних и тех же мотонейронов спинного мозга. Установив этот факт, Шеррингтон охарактеризовал мотонейроны как «общий конечный путь», связывающий головной мозг с мышечным актом. Он изучал движение мышц у животных после перерезки спинного мозга, т.е. после нарушения связи мотонейронов с головным мозгом.

Головной мозг макака, вид сзади. Сектор, величиной 90°, удален, чтобы показать некоторые детали подкорковых структур. Цветная пунктирная линия окружает базальные ганглии левого полушария - части стриатума,скорлупу и хвостатое ядро, а также смежный с ними бледный шар. Ближе к средней линии лежит левый таламус, а по обе стороны от средней линии — правая половина мозжечка, так что видны его внутренние части - зубчатое ядро и ядро шатра. Моторная область коры больших полушарий (окрашена) - филогенетически более поздняя структура по сравнению с базальными ганглиями и мозжечком. Проводимые в настоящее время исследования показывают, что импульсация нейронов моторной коры вызывается сигналами, приходящими в нее через таламус от филогенетически более древних подкорковых структур.
Шеррингтон установил, что через несколько месяцев после такой перерезки у собаки удавались вызвать чесательный рефлекс механическими стимулами - щекотанием кожи или легким потягиванием за волосок где-нибудь на обширной поверхности спины. Описывая эти реакции, он указал, что такие движения «происходили без видимого нарушения направления или ритма». Работа Шеррингтона по чесательному рефлексу привела к современной концепции о «запускаемом движении», основанной на представлении о «центральной программе» с участием спинального генератора ритма. Вскоре после Шеррингтона другой английский физиолог Г. Браун (G. Brown) показал, что у собак, лишенных связей между головным и спинным мозгом, возможны также ритмические движения конечностей, подобные тем, какие происходят при ходьбе. Очевидно, для ходьбы, так же как и для чесания, существуют спинальные генераторы ритма.
Многие теперешние исследования по нейрофизиологии локомоции направлены на выяснение взаимодействия между тем, что можно назвать центральными программами, исходящими от головного мозга, и сенсорными обратными связями. Действительно, работа Шеррингтона особенно касалась того, какими способами активность мотонейронов регулируется сенсорными обратными связями. Он ввел термин «проприоцепция» для обозначения сенсорных входов, которые возникают в процессе вызываемых из центра движений, когда «стимулы, действующие нарецепторы, возникают в самом организме». Шеррингтон избрал приставку «проприо» (от латинского «proprius - «свой собственный»), так как он считал, что главная функция проприоцепторов состоит в том, чтобы по механизму обратной связи давать информацию о собственных движениях организма.
Мышечные проприоцепторы делятся на два вида. Один вид реагирует на удлинение, другой - на напряжение. Мышечные рецепторы длины посылают волокна в спинной мозг и образуют тамсинапсы на мотонейронах, которые оканчиваются на тех же мышцах. Поэтому любое усиление активности рецептора длины, происходящее при удлинении мышцы, активирует мотонейроны удлиненной мышцы. А это вызывает ее сокращение, противодействующее удлинению.
Рецепторы напряжения - второй вид проприоцепторов - чувствительны не к удлинению, а к силе; их активация ведет к торможению связанных с ними мотонейронов. Таким образом, когда прирост напряжения мышцы активирует эти рецепторы, в ответ они действуют на связанные с ними мотонейроны, и это приводит к снижению силы. Поэтому как рецепторы длины, так ирецепторы напряжения можно считать тем, что инженер назвал бы системой управления с помощью отрицательной обратной связи. Данная система поддерживает свою стабильность, противодействуя изменениям длины и напряжения мышцы.
Работа такого сервомеханизма с отрицательной обратной связью станет яснее, если рассмотреть в качестве примера какую-либо такую систему в действии. Представим себе человека, который пытается без наличия внешних помех удерживать свою руку неподвижно вытянутой в сторону. Разумеется, рука при этом непроизвольно слегка колеблется, особенно когда устанет. Например, случайное невольное ослабление напряжения в мышцах, сопротивляющихся силе тяжести, приведет к удлинению этих мышц. А вследствие этого усилится активность одних проприоцепторов -рецепторов длины - и в то же время (вследствие снижения напряжения в мышце) уменьшится активность других проприоцепторов - рецепторов напряжения.

Подопытные обезьяны оказались способными обучаться точным мышечным реакциям на стимулы. В этом опыте экспериментатор зажигал одну из девяти ламп в верхнем ряду панели. Поворотом рукоятки обезьяна могла перемещать верхнюю лампу влево или вправо и получала вознаграждение, когда обе лампы оказывались на одной линии. Микроэлектрод, вживленный в область моторной коры, связанную с точными манипуляциями, регистрировал активность клеток, участвующих в мышечной реакции. Даже незначительное произвольное движение сопровождалось резким усилением активности моторной коры; доля разряжающихся клеток головного мозга была гораздо больше доли активных мотонейронов в спинном мозгу.
Несмотря на то что эти изменения идут в противоположных направлениях, их центральные эффекты не вычитаются один из другого, а суммируются: усиленная импульсация рецепторов длины возбуждает мотонейроны, действующие на мышцы, а ослабление импульсации рецепторов напряжения снимает торможение с тех же самых нейронов. Это синергическое действие двух видов проприоцепторов вступает в силу, когда изменения длины мышцы происходят вследствие непроизвольных изменений в ее напряжении, вызываемых внутренними причинами, но оно не возникнет, если изменения длины и напряжения произойдут вследствие приложения или удаления внешней силы. Например, удлинение мышцы, вызванное увеличением внешней нагрузки, приводит к усилению активности рецептора длины, связанному не с понижением, а с повышением активности рецепторов напряжения. Создавая термин «проприоцептор», Шеррингтон привлек внимание к существенным различиям в нейронной организации, лежащей в основе активного и пассивного движений. В данном контексте «активные» означает собственные движения испытуемого, а «пассивные» — перемещения, производимые внешними силами.

Опыт с «подготовкой к движению» требовал дополнительного обучения обезьян. Их приучали удерживать рукоятку в фиксированном «нейтральном» положении и быть готовыми толкнуть ее вперед через некоторое неопределенное время после вспыхивания зеленого светового сигнала (А) или притянуть назад через некоторое время после вспыхивания красного сигнала (Б). Активность одного нейрона в моторной коре регистрировалась в течение одной секунды до и после сигнальных вспышек; его импульсы записывались в виде горизонтальных рядов точек, где каждый ряд соответствует одному предъявлению стимула. Как показывают эти растровые записи, подготовка к толканию рукоятки вперед вызывала усиление нейронной активности, а подготовка к притягиванию ее вызывала ослабление активности.
Концепция Шеррингтона о связи между рецепторами длины в мышце и движением вообще получила яркое воплощение, когда шведский физиолог Л. Лекселл (L. Leksell) обнаружил, какую роль играют специальные нейроны, называемые гамма-мотонейронами. В отличие от обычных, или альфа-мотонейронов, которые действуют на мышечные волокна, производящие мощное сокращение, гамма-мотонейроны действуют на особые мелкие мышечные волокна, которые регулируют чувствительность рецепторов длины. Следовательно, мотонейроны также бывают двух видов. Один вид действует на мышцы, производящие движения тела (альфа), а второй служит для оптимизации работы рецепторов длины (гамма).
Итак, разные виды мышц, разные виды мотонейронов и связанные с ними системы управления образуют элементарные компоненты механизмов, осуществляющих произвольные и рефлекторные движения. Соотношения между этими компонентами можно изучать в лаборатории на самых разнообразных животных. Например, многие исследования управления движением производятся теперь на моллюсках, а также на членистоногих - насекомых и ракообразных. Простота нервной системы у этих беспозвоночных представляет для исследователя заметные выгоды. Однако эти животные лишены коры большого мозга и связанных с ней структур. Для того чтобы узнать, как головной мозг посылает сигналы мотонейронам и таким образом управляет движениями человека, нужны данные, получаемые на животных, которые обладают не только корой большого мозга, но и ее специализированным отделом — моторной корой, управляющей движением.
Моторная кора была открыта в 1870 г., когда обнаружили, что электрическим раздражением коры большого мозга можно вызвать движения тела. Опыты с такой стимуляцией подтвердили вывод, к которому уже до этого пришел английский невропатолог Дж. Джексон (J. Jackson) на основании клинических наблюдений. Он заметил, что раздражающий очаг повреждения в коре одного полушария большого мозга может вызывать эпилептические движения противоположной стороны тела. Результаты первых опытов по стимуляции, произведенных на собаках, были подтверждены в 1873 г. английским невропатологом Д. Ферье (D. Ferrier) на обезьянах.
Эти исследования моторной коры оказали большое влияние на неврологическое мышление. Чтобы оценить это полностью, надо учесть, что до 1870 г. думали, будто кора больших полушарий служит только для мышления. Джексон выразил тогдашнее общее мнение в следующих словах: «Идея о том, что большие полушария служат для движения, видимо, встречает непреодолимые возражения... Причина этого, как я думаю, лежит в том, что по существующему представлению, извилины [коры] существуют не для движений, а для мыслей».
Следующий шаг был сделан в 1874 г. Он состоял в открытии особой группы гигантских нейронов, которые образуют проводящий путь между моторной корой и спинным мозгом. В этом году русский анатом В. Бец обнаружил в двигательной коре обезьяны и человека необычайно крупные нейроны. Теперь их называют клетками Беца. Было установлено, что аксоны этих клеток спускаются по головному мозгу и образуют прямые связи со спинномозговыми мотонейронами, в частности с нейронами, управляющими теми мышцами, которые служат человеку для точных движений пальцев и речи. Дальнейшее исследование показало, что, казалось бы, непропорционально большая часть моторной коры управляет очень малой долей всей мускулатуры человеческого тела; это наглядно показано на знаменитых «гомункулусах» - картах моторной коры, составленных У. Пенфилдом (W. Penfield) и его сотрудниками из Монреальского неврологического института. Теперь найдены некоторые прямые связи между нейронами моторной коры и мотонейронами грудного отдела спинного мозга, где множество мотонейронов приводит в действие межреберные дыхательные мышцы.
На первый взгляд кажется странным, что нейроны моторной коры, управляющие главным образом точными движениями, оканчиваются на мотонейронах, которые контролируют такое автоматическое и примитивное действие, как дыхание. Но, как указывает английский физиолог Ч. Филиппе (Ch. Phillips), эти связи головного мозга со спинным, вероятно, имеют отношение не к дыханию, а к использованию дыхательных мышц в таких искусных актах, как речь и пение. Таким образом, проекция от моторной коры создает новую регуляцию мышц, предназначенных для старых рефлекторных актов. Как установил сто лет назад Джексон, сама по себе утрата кортико-спинальных связей не парализует мышцы, она препятствует использованию их в некоторых движениях. В частности, разрушение кортико-спинальных путей к грудным мотонейронам не сказывается на использовании дыхательных мышц для дыхания, хотя эти мышцы становятся непригодными для речи.
За последнее десятилетие многое стало известно о контроле над произвольным движением со стороны моторной коры. Новые данные получены в значительной степени благодаря хитроумным методикам, позволяющим вводить микроэлектроды в головной мозг подопытного животного (обычно обезьяны), способного выполнять искусные движения. Пользуясь этими методиками, мы с К. Фроммом (С. Fromm) из Дюссельдорфского университета провели в Национальном институте охраны психического здоровья исследование свойств двигательной коры, лежащих в основе ее решающей роли в точном управлении движениями кисти. Это того же рода точное управление, какое дает возможность хирургу, глядя в микроскоп, передвигать хирургический инструмент с точностью до малых долей миллиметра.
Нам с Фроммом казалось, что если точные мелкие движения управляются выходными сигналами из моторной коры, то тогда ее нейроны должны сильно модулироваться даже мельчайшими флуктуациями мышечной активности. Кроме того, точное управление кистью очень зависит от сенсорной обратной связи, и поэтому мы склонялись к той точке зрения, что активность моторной коры при точно контролируемых движениях кисти должна быть под непрерывным контролем отрицательной обратной связи по замкнутой петле.

Опыт со «стартом», продолжение опыта с подготовкой к движению, имел цель рассмотреть соотношение между произвольными и рефлекторными реакциями. «Стартовый сигнал», производимое мотором перемещение рукоятки из ее нейтрального положения, возникал через одну-пять секунд после включения светового сигнала к подготовке. Активность нейрона моторной коры регистрировали в течение секунды до и после стартового сигнала. Вызывало ли движение рукоятки реакцию толкания (А) или же притягивания к себе (Б), немедленная реакция обезьяны была чисто рефлекторной, о чем на растровых записях говорит усиление нейронной активности сразу же после стимула (через 0,5 секунды). Через 40 миллисекунд этот рефлекторный ответ сменялся произвольным «молчанием» нейрона, связанным с подготовкой к притягиванию рукоятки (белый уча» сток на нижнем растре). После короткого рефлекторного ответа и более продолжительной произвольной реакции (толкания), видных на верхнем растре, животное возвращало рукоятку в нейтральное положение перед следующей пробой; для этого нужно было потянуть ее, что создавало зону молчания, которая видна на верхнем растре (с правой стороны). Точно так же толкание рукоятки для возвращения ее в нейтральное положение вызывало возобновление нейронной активности, видимое на нижнем растре.
Для проверки наших предположений мы начали с того, что обучали обезьян точно передвигать рукоятку прибора. Вращение рукоятки управляло панелью со световыми сигналами; за точные движения обезьяна получала вознаграждение. При каждом тесте регистрировалась импульсация клеток Беца в моторной коре животного. Мы установили, что малейшее перемещение рукоятки сопровождалось поразительным усилением активности этих клеток. Так, доля нейронов моторной коры, которые разряжались при управлении этими тонкими движениями, была гораздо больше доли участвующих в них спинальных мотонейронов.
Была отмечена еще одна особенность в основе той роли, какую играет моторная кора в управлении точными движениями. Это система отрицательной обратной связи, автоматически регулирующая выходную активность моторной коры. Система была сосредоточена преимущественно на тех корковых нейронах, которые контролируют самые точные мелкие движения. Анатомические пути возврата в двигательную кору в системе отрицательной обратной связи детально еще не прослежены. Во всяком случае, одним источником сигналов является соматосенсорная область коры, лежащая непосредственно позади моторной и соединенная с ней множеством связей. Участки соматосенсорной коры, получающие сигналы от кисти, очевидно передают сигналы в моторную кору, замыкая петлю (хотя, вероятно, не единственную) в этой системе обратной связи.
Осуществляемый моторной корой контроль, о котором шла речь выше, имеет решающее значение для возникновения и стабилизации самых точных движений человека. Что происходит, когда человек хочет совершить движение, которое направлено против нормальной рефлекторной реакции? Для исследования этого вопроса мы с Дз. Тандзи (J. Tanji) из Университета Хоккайдо изучали активность нейронов двигательной коры у обезьян, которые были обучены реагировать на непроизвольное движение своей руки. Это обучение иногда требовало мышечной реакции, полностью противоположной нормальному рефлекторному движению. Воспользуемся аналогией с человеком и представим себе испытуемого, который стоит прямо. Он получил инструкцию: когда экспериментатор толкает его, он должен наклониться вперед, не переставляя ног. Испытуемый сначала подготовится к движению вперед, а затем будет ждать толчка. Если толчок последует сзади, то для сохранения равновесия испытуемый рефлекторно отогнется назад. Для осуществления центрально запрограммированного движения, направленного вперед в соответствии с инструкцией, он должен теперь выйти из способа реагирования по замкнутой петле, который сохраняет ему равновесие, и перейти к реакции по открытой петле, которая приведет к тому, что он качнется вперед (в данном случае упадет в сетку или на руки к тому, кто стоит наготове, чтобы поймать его).
Мы с Тандзи обучали обезьян отвечать на такую же последовательность событий. Животные начинали с того, что точно устанавливали рукоятку и удерживали ее в неподвижности в течение нескольких секунд. Во время этого короткого периода выход сигналов из их моторной коры к мышцам руки регулировался отрицательной обратной связью по замкнутой петле. Затем включалась цветная лампа. Ее цвет показывал обезьяне, как реагировать на предстоящее перемещение рукоятки извне. Если свет был красный, обезьяна должна была тянуть рукоятку назад, если зеленый - толкать вперед. Обезьяны получали вознаграждение за правильные ответы на движения рукояток после получения обоих сигналов. Подготовка к реакции на эти сигналы занимала около 200 миллисекунд.
Регистрируя импульсацию нейронов моторной коры, мы отметили, что после подготовки к движению животному нужно было всего 40 миллисекунд, чтобы правильно реагировать на движения рукоятки. В коротком интервале, следующем за движением, корковый контроль резко переходил от обратной связи по замкнутой петле (которая рефлекторно обеспечивает устойчивость позы) к связи по открытой петле, которая была нужна, чтобы совершить запрограммированное движение.
Итак, соматосенсорная область коры больших полушарий выполняет функцию передачи сигналов, контролирующих выходную активность моторной коры по принципу замкнутой петли. Но соматосенсорная область не посылает тех сигналов, которые лежат в основе запрограммированных движений, осуществляемых по принципу открытой петли, несмотря на (а не вследствие) рефлекторные эффекты соматосенсорной входной активности. Сигналы, связанные с запрограммированными движениями, приходят в двигательную кору из подкорковых структур, в особенности из мозжечка через таламус. П. Стрик (P. Strick) из Национальных институтов здравоохранения поставил опыты, которые показали, что в центрально запрограммированном управлении по открытой петле участвует путь, идущий от мозжечка через таламус в кору. Проводя опыты в основном по тому же плану, что и мы с Тандзи, Стрик регистрировал активность отдельных нейронов в определенных структурах мозжечка.
Он обучал своих обезьян двигать рукой в ответ на поданный знак, состоявший в перемещении руки посредством производимого извне передвижения рукоятки, которую держала обезьяна. Стрик обнаружил, что если предварительно у обезьян при помощи системы красных и зеленых сигналов было запрограммировано перемещение рукоятки в том или другом направлении, то эта программа оказывала сильное влияние па реакции определенных, так называемых зубчатых нейронов мозжечка: эти нейроны разряжались в пределах 30 миллисекунд после того, как был подан знак в виде перемещения руки. Тем самым у активности зубчатого нейрона было 10 миллисекунд на то, чтобы пройти через таламус и вызвать, центрально запрограммированную активность моторной коры (проявляющуюся через 40 миллисекунд после подачи знака).

Проводящие пути между некоторыми областями коры больших полушарий и определенными подкорковыми структурами показаны на схеме продольного разреза большого мозга обезьяны. Тонкими стрелками обозначены входы в базальные ганглии (структуры внутри черных границ), которые проводят разнообразную информацию от коры больших полушарий. Один из компонентов, стриатум, является главным связующим звеном между ассоциативными областями коры и моторной областью. Путь, выходящий из стриатума, проходит к разделенному на две части бледному шару (толстая стрелка); это образование в свою очередь посылает связи в другую подкорковую структуру - таламус, в особенности к двум его ядрам: ventralis lateralis и ventralis anterior (толстая и менее толстая стрелки). Пути, выходящие из таламуса, идут главным образом в премоторную область коры (толстая стрелка); дополнительные пути (менее толстая стрелка) направляются в моторную кору, откуда сигналы идут к мотонейронам спинного мозга (пунктирная стрелка). Подкорковые входы играют важную роль в центральном программировании движений.
Факты, полученные Стриком, согласуются с данными У. Тэтча мл. (W. Thatch, Jr.) из Вашингтонского университета. Работая в Национальном институте охраны психического здоровья, Тэтч показал, что импульсация нейронов мозжечка намного опережала мышечную активность у обезьяны, обученной отвечать на световой стимул. Роль сигналов, идущих от мозжечка, в генерации активности моторной коры была также экспериментально показана В. Бруксом (V. Brooks) с сотрудниками из Университета Западного Онтарио. Они искусственно понижали температуру мозжечка у обезьян, а затем давали ей вернуться к норме. Активность нейронов двигательной коры измерялась до, во время и после охлаждения мозжечка. Было установлено, что во время охлаждения импульсация нейронов моторной коры и соответствующее запрограммированное движение запаздывали.
Кроме проведения сигналов от мозжечка в моторную кору таламус передает сигналы еще от одной подкорковой структуры, а именно от большой совокупности клеточных групп, объединяемых общим названием базальных ганглиев. Во время своей работы в Национальном институте охраны психического здоровья М. Де-Лонг (М. DeLong) из Университета Джонса Гопкинса показал, что клетки базальных ганглиев разряжаются задолго до произвольных движений, совершаемых животным в ответ на сигналы, Этот факт согласуется с результатами наблюдений, сделанных в неврологических отделениях; согласно этим наблюдениям, базальные ганглии имеют решающее значение для самых ранних стадий инициации движения - стадий, когда в результате еще не разгаданных процессов абстрактная мысль переводится в конкретный двигательный акт.
Рефлексы и произвольные движения не противоположны друг другу. Это признал еще сто лет назад Джексон, когда писал, что произвольные движения подчиняются законам, управляющим рефлекторными актами. Однако если произвольным движениям нельзя дать определение путем исключения, т. е. определить их как нечто такое, что не является рефлексом, то в таком случае как же их определить? Самое сжатое из известных мне определений дал шведский нейрофизиолог Р. Гранит (R. Granit) в своей недавно опубликованной книге «Целенаправленный мозг»: «Произвольным в произвольном движении является его цель». С такой точки зрения произвольные свойства моторного акта надо рассматривать в аспекте цели совершаемого действия. В то же время действительные события, лежащие в основе достижения цели, строятся из разнообразных рефлекторных процессов.
Недавно я обсуждал этот вопрос с русским кибернетиком Виктором Гурфинкелем. Он тоже определяет произвольное движение в связи с его целью. Описывая виды процессов, на которых основано произвольное движение, он рассказал мне о некоторых кинезиологических исследованиях, направленных на оценку характеристик управления движением у лучших стрелков-спортсменов. Гурфинкель указал, что важным свойством меткого стрелка является его способность удерживать пистолет неподвижным. Как показали исследования электромиографических и кинематических характеристик снайперов, при том что многие части их тела двигались, пистолет оставался практически неподви